
Биосинтез компонентов вируса характеристика
Типы взаимодействия вируса с клеткой. Основные стадии репродукции вирусов. Прикрепление вирусов к поверхности клетки (адсорбция вирусов). Способы проникновения вирусов животных в клетку. Биосинтез компонентов вируса. Формирование (сборка) вирусов.
| Рубрика | Сельское, лесное хозяйство и землепользование |
| Вид | реферат |
| Язык | русский |
| Дата добавления | 11.10.2016 |
| Размер файла | 38,4 K |
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
Систематика, морфология, антигенные свойства. Патогенность, место репродукции, восприимчивые животные, лабораторные модели. Устойчивость вируса. Характеристика болезни вызываемой вирусом. Определение(синономы). Эпизоотологические данные. Патогенез.
контрольная работа [18,3 K], добавлен 06.11.2007
Специфические факторы противовирусного иммунитета. Два варианта выдачи иммунного ответа в форме биосинтеза антител. Вирус инфекционного бронхита птиц: возбудитель, диагностика. Методы лечения вируса ящура. Культивирование вирусов в культуре клеток.
курсовая работа [49,2 K], добавлен 17.11.2010
Понятие, сущность, типы, динамика и способы распространения эпифитотия, а также роль патогена, растения-хозяина и окружающей среды в его развитии. Анализ путей передачи вирусов растений. Описание мер борьбы, по защите растений от инфекционных заболеваний.
реферат [23,5 K], добавлен 14.11.2010
Роль условно-патогенных бактерий и вирусов в этиопатогенезе острых кишечных и респираторных болезней. Применение для профилактики и лечения специфических поливалентных вакцин и сывороток крови. Пути повышения резистентности сельскохозяйственных животных.
курсовая работа [70,3 K], добавлен 05.01.2011
Негативные последствия болезней, вызываемых слабопатогенными вирусами. Методы выделения вирусов из материала больных животных и трупов. Возбудитель и эпизоотология оспы птиц, ее профилактика и лечение. Клинические признаки и диагностика бешенства у коров.
контрольная работа [44,2 K], добавлен 23.10.2013
Уравновешивание популяции вредителей. Основные особенности функционирования экологических систем. Биологическая борьба с вредными видами организмов. Численность популяций отдельных видов. Охрана полезных организмов и вирусов и их массовая интродукция.
реферат [254,5 K], добавлен 21.07.2011
Характеристика понятия эпифитотии. Ознакомление с путями передачи вирусов от одного растения к другому. Рассмотрение симптом местных, прогрессирующих и повсеместных эпифитотий. Описание основных методов защиты растений от инфекционных заболеваний.
презентация [735,3 K], добавлен 07.11.2013
Таксономия, этапы репродукции вируса ринотрахеита кошек. Основной путь заражения. Особенности культивирования в различных живых системах. Клинические признаки заболевания. Принципы диагностики герпес-вирусной инфекции методом полимеразной цепной реакции.
реферат [1,6 M], добавлен 02.06.2015
Определение и история открытия заболевания. Этиология вируса африканской чумы свиней. Эпизоотология, клинические признаки и патогенез. Основные методы выделения вируса и выявления антигенов. Патологоанатомические изменения, дифференциальная диагностика.
курсовая работа [10,1 M], добавлен 20.11.2013
Таксономия вируса африканской чумы свиней, характеристика вириона, распространение, степень опасности и ущерб. Антигенные свойства вируса АЧС. Гемадсорбирующая активность и культуральные свойства. Этапы лабораторной диагностики и методы профилактики.
реферат [244,2 K], добавлен 20.12.2016

Работы в архивах красиво оформлены согласно требованиям ВУЗов и содержат рисунки, диаграммы, формулы и т.д.
PPT, PPTX и PDF-файлы представлены только в архивах.
Рекомендуем скачать работу.
1.у вируса оспы линейная двуцепочечная ДНК, синтез идет как обычно;2.у ВТМ одноцепочечная +РНК. В ней закодирован белок капсида, а так же фермент РНК-зависимая РНК-полимераза, которая на +РНК строит -РНК, а затем на ней, как на матрице, делает множество +РНК;3.у гриппа РНК-зависимая РНК-полимераза находится внутри вириона. Однонитчатая -РНК гриппа состоит из нескольких фрагментов, синтез единой плюс-РНК происходит внутри ядра клетки, включает в себя сплайсинг. Такой механизм обеспечивает очень высокий уровень генетической рекомбинации, в том числе и между разными штаммами вируса.
Процесс формирования зрелых вирионов носит следующий характер. Синтезированные, функционально активные вирионные полипептиды НА+ и NA+ транспортируются к наружной плазматической мембране и на определенном участке внедряются в ее структуру, вытесняя клеточные полипептиды [Харитоненков И. Т., 1980; Lubeck M. et al., 1979].
На этом участке клеточной поверхности появляются выступы (шипы), характерные для поверхности зрелого вириона. Вирионные гликопротеиды в отличие от клеточных не проникают глубоко в липидный слой и лишены способности мигрировать на поверхности мембраны. Низкая подвижность НА+ и NA+ может быть связана с их прочным взаимодействием с молекулами М-белка, которые также транспортируются к участку модифицированной мембраны. М-белок, вытесняя клеточные негликозилированные полипептиды, образует внутреннюю выстилку липидной мембраны. На следующем этапе к сформировавшемуся участку будущей вирусной мембраны транспортируются вирионные РНП, тесно связанные с белками полимеразного комплекса. Модифицированная мембрана окружает РНП, ее структура начинает выпячиваться, М-белок формирует нуклеокапсид, окружая РНП и белки полимеразного комплекса, и вирион отпочковывается от поверхности зараженной клетки. На протяжении всего процесса вирусного созревания плазматическая мембрана и ее модифицированный участок неразрывно связаны между собой, образуя непрерывную структурную единицу.
Реализация генетической информации вируса гриппа осуществляется с помощью структур и механизмов, обеспечивающих в норме биосинтетические потребности клетки-хозяина. Поэтому взаимодействие вируса и клетки при продуктивном способе репродукции сопровождается более или менее выраженными изменениями метаболизма хозяина, определяющими экспрессию вирусного генома и регулирующими биосинтез вирусного потомства. Стадия пенетрации вируса и освобождения функционально активного генома из окружающих его оболочек происходит в чувствительной клетке на фоне отчетливой активации эндогенного дыхания [Мещерякова И. Е. и др., 1976], усиленного фосфорилирования мембранных белков, активации циклазной системы и микротабулярных процессов [Krizanova О. et al., 1977]. Это обеспечивает энергетические расходы клетки-хозяина на осуществление энергозависимых этапов репродукции вируса гриппа [Поляк Р. Я., Дубровина Т. Я., 1980].
Типичная фаговая частица (вирион) состоит из головки и хвоста. Длина хвоста обычно в 2 — 4 раза больше диаметра головки. В головке содержится генетический материал — одноцепочечная или двуцепочечная РНК или ДНК с ферментом транскриптазой в неактивном состоянии, окруженная белковой или липопротеиновой оболочкой — капсидом, сохраняющим геном вне клетки [9] .Нуклеиновая кислота и капсид вместе составляют нуклеокапсид. Бактериофаги могут иметь икосаэдральный капсид, собранный из множества копий одного или двух специфичных белков. Хвост представляет собой белковую трубку — продолжение белковой оболочки головки, в основании хвоста имеется АТФаза, которая регенерирует энергию для инъекции генетического материала. Фаги, как и все вирусы, являются абсолютными внутриклеточными паразитами. Хотя они переносят всю информацию для запуска собственной репродукции в соответствующем хозяине, у них отсутствуют механизмы для выработки энергии и рибосомы для синтеза белка.
Под резистентностью (устойчивостью) понимают способность микроорганизма переносить значительно большие концентрации препарата, чем остальные микроорганизмы данного штамма (вида), или развиваться при таких концентрациях, которые превышают достигаемые в макроорганизме при введении антибиотиков, сульфаниламидов и нитрофуранов в терапевтических дозах. Резистентные штаммы микроорганизмов возникают при изменении генома бактериальной клетки в результате спонтанных мутаций. Последние не связаны с направленным действием на ДНК бактерии антибактериальных препаратов, которые играют роль лишь селективных агентов. В процессе селекции в результате воздействия химиотерапевтических соединений чувствительные микроорганизмы погибают, а резистентные сохраняются, размножаются и распространяются в окружающей среде. Приобретенная резистентность закрепляется и передается по наследству последующим генерациям бактерий.
Биологическое явление симбиоза микробной клетки с умеренным фагом (профагом) называется лизогенией, а культура бактерий, содержащая профаг, получила название лизогенной. Это название отражает способность профага самопроизвольно или под действием ряда физических и химических факторов исключаться из хромосомы клетки и переходить в цитоплазму, т. е. вести себя как вирулентный фаг, лизирующий бактерии. Лизогенные культуры по своим основным свойствам не отличаются от исходных, но они невосприимчивы к повторному заражению гомологичным или близкородственным фагом и, кроме того, приобретают дополнительные свойства, которые находятся под контролем генов профага. Изменение свойств микроорганизмов под влиянием профага получило название фаговой конверсии. Последняя имеет место у многих видов микроорганизмов и касается различных их свойств: культуральных, биохимических, токсигенных, антигенных, чувствительности к антибиотикам и др. Кроме того, переходя из интегрированного состояния в вирулентную форму, умеренный фаг может захватить часть хромосомы клетки и при лизисе последней переносит эту часть хромосомы в другую клетку. Если микробная клетка станет лизогенной, она приобретает новые свойства. Таким образом, умеренные фаги являются мощным фактором изменчивости микроорганизмов.
Лизогенная конверсия(синоним фаговая конверсия) — изменение свойств бактериальной клетки вследствие заражения ее умеренным бактериофагом. Лизогенная конверсия связана с добавлением к бактериальному геному новой генетической информации, которая вносится в клетку бактерии геномом фага. Она может происходить также путем трансдукции и являться причиной подавления или усиления ферментативной активности, изменения патогенных свойств, морфологии колоний, устойчивости к антибиотикам и др.
Вскоре после обнажения вирусного генома происходит уменьшение или исчезновение инфекционности, поскольку вирионов как таковых уже не существует, а инфекционность нуклеиновой кислоты намного меньше инфекционности полных вирионов. Данное явление было названо эклипсом (затмением), а фаза вирусной инфекции с момента адсорбции и до появления в клетке новых инфекционных вирионов — эклипс-фазой, или латентным периодом. При высокой множественности заражения клеток некоторая часть вирионов сохраняет инфекционные свойства в течение всей эклипс-фазы, что зачастую затрудняет определение ее продолжительности.
В течение этого периода происходят все существенные процессы вирусной репродукции, приводящие к образованию вирусных компонентов и формированию нового потомства вирионов. С появлением в клетке первых вновь синтезированных вирионов завершается эклипс-период размножения вируса.
Процессы, происходящие в эклипс-фазе, схематично показаны на рисунке, где в качестве примера представлены основные этапы репликации аденовирусов.
Из представленной схемы видно, что после прикрепления вирион входит в клетку и частично или полностью лишается оболочки, обнажая вирусный геном. Некоторые ранние вирусные гены транскрибируются с образованием мРНК, которые могут затем изменяться различными путями, включая сплайсинг. Ранние мРНК транслируются с образованием трех главных продуктов: белков, которые подавляют или останавливают синтез белков и нуклеиновых кислот; белков, которые регулируют экспрессию вирусного генома; ферментов, необходимых для репликации вирусных нуклеиновых кислот. Вслед за репликацией вирусной нуклеиновой кислоты транскрибируются поздние вирусные гены, продуктами трансляции которых являются в основном структурные вирусные белки, используемые при сборке новых вирионов. Некоторые из них претерпевают посттрансляционные изменения перед включением в структуру вирионов. В каждой инфицированной клетке образуются сотни и тысячи вирионов, которые, освобождаясь, заражают другие клетки.
У большинства ДНК-вирусов транскрипция и репликация ДНК происходит в ядре клетки, с использованием клеточной РНК-полимеразы II и других клеточных ферментов. Большинство РНК-вирусов размножается в цитоплазме, где отсутствуют ферменты, копирующие РНК на РНК-матрице, поэтому вирусный геном должен сам по себе функционировать как мРНК или вирус должен нести свою РНК-полимеразу, чтобы транскрибировать РНК на РНК-геноме.
Синтез вирусных белков происходит только в цитоплазме клетки. В инфицированных клетках вирусные нуклеиновые кислоты и вирусспецифические белки синтезируются в значительно большем количестве, чем включаются в вирионы. Избыточный синтез структурных компонентов при репродукции вирусов является своеобразной платой за паразитизм на генетическом уровне.
Потомство вирионов проявляется спустя определенный период после того, как начал осуществляться синтез составляющих их компонентов. Продолжительность периода от момента инфицирования до появления дочерних вирионов довольно вариабельна и отражает глубокое различие между разными вирусами.
Латентный период мелких вирусов, не имеющих оболочки и содержащих односпиральную РНК, длится примерно 2 часа и является наиболее коротким среди всех известных вирусов животных. ДНК-содержащие вирусы имеют более продолжительный латентный период. Длительный латентный период отмечен также у реовирусов и вируса саркомы Рауса. У простых мелких РНК-содержащих вирусов быстрее протекает не только латентный период, но и весь цикл размножения вируса. Вирусы, геном которых выполняет функции мРНК, имеют более короткий репликативный цикл, чем вирусы, у которых функции мРНК выполняет не вирионная РНК, а комплементарная ей минус-нить.
Размножение ДНК-содержащих вирусов может продолжаться даже через 24 часа после заражения. Причины, определяющие длительность цикла размножения разных вирусов, не выяснены. Возможно, что это связано с более сложной последовательностью событий при репродукции крупных, сложно устроенных вирусов.
Укорочение латентного периода при высокой множественности заражения было обнаружено у различных РНК и ДНК вирусов и, возможно, связано с ускорением процесса образования ферментов, участвующих в синтезе нуклеиновой кислоты. При высокой множественности заражения не исключается также роль капсидных белков в ускорении начала цикла репродукции.
Число инфекционных вирусных частиц, образуемых в одной клетке, зависит от типа вируса, и количество их варьирует очень широко. Интересно, что на долю вирусспецифических продуктов приходится от 0,1 до 5% массы клетки животного.
Вирусы — мельчайшие микробы, не имеющие клеточного строения, белоксинтезирующей системы, содержащие только ДНК или РНК. Относятся к царству Vira. Являясь облигатными внутриклеточными паразитами, вирусы размножаются в цитоплазме или ядре клетки. Они — автономные генетические структуры. Отличаются особым — разобщенным (дисъюнктивным) способом размножения (репродукции): в клетке отдельно синтезируются нуклеиновые кислоты вирусов и их белки, затем происходит их сборка в вирусные частицы. Сформированная вирусная частица называется вирионом.
Морфологию и структуру вирусов изучают с помощью электронного микроскопа, так как их размеры малы и сравнимы с толщиной оболочки бактерий.
Форма вирионов может быть различной: палочковидной (вирус табачной мозаики), пулевидной (вирус бешенства), сферической (вирусы полиомиелита, ВИЧ), в виде сперматозоида (многие бактериофаги). Различают просто устроенные и сложно устроенные вирусы.
Простые, или безоболочечные, вирусы состоят из нуклеиновой кислоты и белковой оболочки, называемой капсидом. Капсид состоит из повторяющихся морфологических субъединиц — капсомеров. Нуклеиновая кислота и капсид взаимодействуют друг с другом, образуя нуклеокапсид.
Сложные, или оболочечные, вирусы снаружи капсида окружены липопротеиновой оболочкой (суперкапсидом, или пеплосом). Эта оболочка является производной структурой от мембран вирус-инфицированной клетки. На оболочке вируса расположены гликопротеиновые шипы, или шипики (пепломеры). Под оболочкой некоторых вирусов находится матриксный М-белок.
Тип симметрии. Капсид или нуклеокапсид могут иметь спиральный, икосаэдрический (кубический) или сложный тип симметрии. Икосаэдрический тип симметрии обусловлен образованием изометрически полого тела из капсида, содержащего вирусную нуклеиновую кислоту (например, у вирусов гепатита А, герпеса, полиомиелита). Спиральный тип симметрии обусловлен винтообразной структурой нуклеокапсида (например, у вируса гриппа).
Включения — скопление вирионов или отдельных их компонентов в цитоплазме или ядре клеток, выявляемые под микроскопом при специальном окрашивании. Вирус натуральной оспы образует цитоплазмати-ческие включения — тельца Гварниери; вирусы герпеса и аденовирусы — внутриядерные включения.
Размеры вирусов определяют с помощью электронной микроскопии, методом ультрафильтрации через фильтры с известным диаметром пор, методом ультрацентрифугирования. Одним из самых мелких вирусов является вирус полиомиелита (около 20 нм), наиболее крупным — натуральной оспы (около 350 нм).
Вирусы имеют уникальный геном, так как содержат либо ДНК, либо РНК. Поэтому различают ДНК-содержащие и РНК-содержащие вирусы. Они обычно гаплоидны, т.е. имеют один набор генов. Геном вирусов представлен различными видами нуклеиновых кислот: двунитчатыми, однонитчатыми, линейными, кольцевыми, фрагментированными. Среди РНК-содержащих вирусов различают вирусы с положительным (плюс-нить РНК) геномом. Плюс-нить РНК этих вирусов выполняет наследственную функцию и функцию информационной РНК (иРНК). Имеются также РНК-содержащие вирусы с отрицательным (минус-нить РНК) геномом. Минус-нить РНК этих вирусов выполняет только наследственную функцию.
Вирусы поражают позвоночных и беспозвоночных животных, а также растения и бактерии. Являясь основными возбудителями инфекционных заболеваний человека, вирусы также участвуют в процессах канцерогенеза, могут передаваться различными путями, в том числе через плаценту (вирус краснухи, цитомегаловирус и др.), поражая плод человека. Они могут приводить к постинфекционным осложнениям — развитию миокардитов, панкреатитов, иммунодефицитов и др.
Кроме обычных вирусов, известны и так называемые неканонические вирусы — прионы — белковые инфекционные частицы, являющиеся агентами белковой природы, имеющие вид фибрилл размером 10—20x100—200 нм. Прионы, по-видимому, являются одновременно индукторами и продуктами автономного гена человека или животного и вызывают у них энцефалопатии в условиях медленной вирусной инфекции (болезни Крейтц-фельдта—Якоба, куру и др.).
Другими необычными агентами, близкими к вирусам, являются вироиды — небольшие молекулы кольцевой, суперспирализованной РНК, не содержащие белка, вызывающие заболевания у растений.
Типы взаимодействия вируса с клеткой. Различают три типа взаимодействия вируса с клеткой: продуктивный, абортивный и интегративный.
Продуктивный тип — завершается образованием нового поколения вирионов и гибелью (лизисом) зараженных клеток (цитолитическая форма). Некоторые вирусы выходят из клеток, не разрушая их (нецитолитическая форма).
Абортивный тип — не завершается образованием новых вирионов, поскольку инфекционный процесс в клетке прерывается на одном из этапов.
Интегративный тип, или вирогения — характеризуется встраиванием (интеграцией) вирусной ДНК в виде провируса в хромосому клетки и их совместным сосуществованием (совместная репликация).
Адсорбция. Взаимодействие вируса с клеткой начинается с процесса адсорбции, т. е. прикрепления вирусов к поверхности клетки. Это высокоспецифический процесс. Вирус адсорбируется на определенных участках клеточной мембраны — так называемых рецепторах. Клеточные рецепторы могут иметь разную химическую природу, представляя собой белки, углеводные компоненты белков и липидов, липиды. Число специфических рецепторов на поверхности одной клетки колеблется от 10 4 до 10 5 . Следовательно, на клетке могут адсорбироваться десятки и даже сотни вирусных частиц.
Проникновение в клетку. Существует два способа проникновения вирусов животных в клетку: виропексис и слияние вирусной оболочки с клеточной мембраной. При виропексисе после адсорбции вирусов происходят инвагинация (впячивание) участка клеточной мембраны и образование внутриклеточной вакуоли, которая содержит вирусную частицу. Вакуоль с вирусом может транспортироваться в любом направлении в разные участки цитоплазмы или ядро клетки. Процесс слияния осуществляется одним из поверхностных вирусных белков капсидной или суперкапсидной оболочки. По-видимому, оба механизма проникновения вируса в клетку не исключают, а дополняют друг друга.
Биосинтез компонентов вируса. Проникшая в клетку вирусная нуклеиновая кислота несет генетическую информацию, которая успешно конкурирует с генетической информацией клетки. Она дезорганизует работу клеточных систем, подавляет собственный метаболизм клетки и заставляет ее синтезировать новые вирусные белки и нуклеиновые кислоты, идущие на построение вирусного потомства.
Реализация генетической информации вируса осуществляется в соответствии с процессами транскрипции, трансляции и репликации.
Существуют следующие общие принципы сборки вирусов, имеющих разную структуру:
1. Формирование вирусов является многоступенчатым процессом с образованием промежуточных форм;
2. Сборка просто устроенных вирусов заключается во взаимодействии молекул вирусных нуклеиновых кислот с капсидными белками и образовании нуклеокапсидов (например, вирусы полиомиелита). У сложно устроенных вирусов сначала формируются нуклеокапсиды, с которыми взаимодействуют белки суперкапсидных оболочек (например, вирусы гриппа);
3. Формирование вирусов происходит не во внутриклеточной жидкости, а на ядерных или цитоплазматических мембранах клетки;
4. Сложно организованные вирусы в процессе формирования включают в свой состав компоненты клетки-хозяина (липиды, углеводы).
Время, необходимое для осуществления полного цикла репродукции вирусов, варьирует от 5—6 ч (вирусы гриппа, натуральной оспы и др.) до нескольких суток (вирусы кори, аденовирусы и др.). Образовавшиеся вирусы способны инфицировать новые клетки и проходить в них указанный выше цикл репродукции.
Структура и химический состав бактериофагов. Взаимодействие фага с бактериальной клеткой. Умеренные и вирулентные бактериофаги. Лизогения. Применение фагов в биотехнологии, микробиологии и медицине.
Бактериофаги — вирусы бактерий, обладающие способностью специфически проникать в бактериальные клетки, репродуцироваться в них и вызывать их растворение (лизис).
В зависимости от формы и структурной организации фаги подразделяют на несколько морфологических типов: нитевидные, мелкие кубические, фаги сперматозоидной формы.
Взаимодействие фага с бактериальной клеткой. По механизму взаимодействия различают вирулентные и умеренные фаги.
После биосинтеза фаговых компонентов и их самосборки в бактериальной клетке накапливается до 200 новых фаговых частиц. Под действием фагового лизоцима и внутриклеточного осмотического давления происходит разрушение клеточной стенки, выход фагового потомства в окружающую среду и лизис бактерии. Один литический цикл (от момента адсорбции фагов до их выхода из клетки) продолжается 30—40 мин. Процесс бактериофагии проходит несколько циклов, пока не будут лизированы все чувствительные к данному фагу бактерии.
Взаимодействие фагов с бактериальной клеткой характеризуется определенной степенью специфичности. По специфичности действия различают поливалентные фаги, способные взаимодействовать с родственными видами бактерий, моновалентные фаги, взаимодействующие с бактериями определенного вида, и типовые фаги, взаимодействующие с отдельными вариантами (типами) данного вида бактерий.
Умеренные фаги лизируют не все клетки в популяции, с частью из них они вступают в симбиоз, в результате чего ДНК фага встраивается в хромосому бактерии. В таком случае геномом фага называют профаг. Профаг, ставший частью хромосомы клетки, при ее размножении реплицируется синхронно с геном бактерии, не вызывая ее лизиса, и передается по наследству от клетки к клетке неограниченному числу потомков.
Биологическое явление симбиоза микробной клетки с умеренным фагом (профагом) называется лизогенией, а культура бактерий, содержащая профаг, получила название лизогенной. Это название отражает способность профага самопроизвольно или под действием ряда физических и химических факторов исключаться из хромосомы клетки и переходить в цитоплазму, т. е. вести себя как вирулентный фаг, лизирующий бактерии.
Лизогенные культуры по своим основным свойствам не отличаются от исходных, но они невосприимчивы к повторному заражению гомологичным или близкородственным фагом и, кроме того, приобретают дополнительные свойства, которые находятся под контролем генов профага. Изменение свойств микроорганизмов под влиянием профага получило название фаговой конверсии. Последняя имеет место у многих видов микроорганизмов и касается различных их свойств: культуральных, биохимических, токсигенных, антигенных, чувствительности к антибиотикам и др. Кроме того, переходя из интегрированного состояния в вирулентную форму, умеренный фаг может захватить часть хромосомы клетки и при лизисе последней переносит эту часть хромосомы в другую клетку. Если микробная клетка станет лизогенной, она приобретает новые свойства. Таким образом, умеренные фаги являются мощным фактором изменчивости микроорганизмов.
По содержанию бактериофагов в объектах окружающей среды (например, в воде) можно судить о присутствии в них соответствующих патогенных бактерий. Подобные исследования проводят при эпидемиологическом анализе вспышек инфекционных болезней.
Фаги применяют также для лечения и профилактики ряда бактериальных инфекций. Производят брюшнотифозный, сальмонеллезный, дизентерийный, синегнойный, стафилококковый, стрептококковый фаги и комбинированные препараты (колипротейный, пиобактериофаги и др). Бактериофаги назначают по показаниям перорально, парентерально или местно в виде жидких, таблети-рованных форм, свечей или аэрозолей.
Бактериофаги широко применяют в генной инженерии и биотехнологии в качестве векторов для получения рекомбинантных ДНК.
Вирусы – это мельчайшие микроорганизмы, относящиеся к царству Vira, не имеющие клеточного строения, белоксинтезирующей системы, содержащие только один тип нуклеиновой кислоты (ДНК или РНК).
Они отличаются особым разобщенным (дисъюнктивным) способом размножения (репродукции): в клетке отдельно синтезируются нуклеиновые кислоты вирусов и их белки и затем происходит их сборка в вирусные частицы. Вирусы, являясь облигатными внутриклеточными паразитами, размножаются в цитоплазме или ядре клетки. Сформированная вирусная частица называется вирионом.
Морфологию и структуру вирусов изучают с помощью электронного микроскопа, так как их размеры малы и сравнимы с толщиной оболочки бактерий.
Вирусы имеют различную форму вирионов:
- палочковидной (вирус табачной мозаики)
- пулевидной (вирус бешенства)
- сферической (вирусы полиомиелита, ВИЧ)
- в виде сперматозоида (многие бактериофаги).
Вирусы имеют разные размеры, которые определяют с помощью электронной микроскопии, методом ультрафильтрации через фильтры с известным диаметром пор, методом ультрацентрифугирования. Одним из самых мелких вирусов является вирус полиомиелита (около 20 нм), наиболее крупным – натуральной оспы (около 350 нм).
Вирусы имеют уникальный геном, так как содержат либо ДНК, либо РНК. Поэтому различают ДНК-содержащие и РНК-содержащие вирусы. Они обычно гаплоидны, т.е. имеют один набор генов. Геном вирусов представлен различными видами нуклеиновых кислот: двунитчатыми, однонитчатыми, линейными, кольцевыми, фрагментированными.
Среди РНК-содержащих вирусов различают вирусы с положительным (плюс-нить РНК) геномом. Плюс-нить РНК этих вирусов выполняет наследственную функцию и функцию информационной РНК (иРНК). Имеются также РНК-содержащие вирусы с отрицательным (минус-нить РНК) геномом.
Минус-нить РНК этих вирусов выполняет только наследственную функцию. Геном вирусов способен включаться в состав генетического аппарата клетки в виде провируса, проявляя себя генетическим паразитом клетки. Нуклеиновые кислоты некоторых вирусов (вирусы герпеса и др.) могут находиться в цитоплазме инфицированных клеток, напоминая плазмиды.
Вирусы различают по строению:
- просто устроенные (например, вирус полиомиелита)
- сложно устроенные (например, вирусы гриппа, кори) вирусы.
У просто устроенных вирусов нуклеиновая кислота связана с белковой оболочкой, называемой капсидом (от лат. capsa – футляр). Капсид состоит из повторяющихся морфологических субъединиц – капсомеров. Нуклеиновая кислота и капсид, взаимодействуя друг с другом, образуют нуклеокапсид.
Вирусы различают по типу симметрии капсида:
- спиральный – обусловлен винтообразной структурой нуклеокапсида
- кубический– обусловлен образованием изометрически полого тела из капсида, содержащего вирусную нуклеиновую кислоту.
- сложный
Капсид и суперкапсид защищают вирионы от влияния окружающей среды, обусловливают избирательное взаимодействие (адсорбцию) с клетками, определяют антигенные и иммуногенные свойства вирионов. Внутренние структуры вирусов называются сердцевиной.
В вирусологии используют следующие таксономические категории:
- семейство (название оканчивается на viridae)
- подсемейство (название оканчивается на virinae)
- род (название оканчивается на virus).
Однако названия родов и особенно подсемейств сформулированы не для всех вирусов. Вид вируса биноминального названия, как у бактерий, не получил.
Вирусы классифицируются по следующим категориям:
- тип нуклеиновой кислоты (ДНК или РНК), ее структура, количество нитей (одна или две),
- особенности воспроизводства вирусного генома;
- размер и морфология вирионов, количество капсомеров и тип симметрии;
- наличие суперкапсида;
- чувствительность к эфиру и дезоксихолату;
- место размножения в клетке;
- антигенные свойства и пр.
Вирусы поражают позвоночных и беспозвоночных животных, а также растения и бактерии. Являясь основными возбудителями инфекционных заболеваний человека, вирусы также участвуют в процессах канцерогенеза, могут передаваться различными путями, в том числе через плаценту (вирус краснухи, цитомегаловирус и др.), поражая плод человека. Они могут приводить к постинфекционным осложнениям – развитию миокардитов, панкреатитов, иммунодефицитов и др.
Кроме обычных вирусов, известны и так называемые неканонические вирусы – прионы – белковые инфекционные частицы, являющиеся агентами белковой природы, имеющие вид фибрилл размером 10.20×100.200 нм. Прионы, по-видимому, являются одновременно индукторами и продуктами автономного гена человека или животного и вызывают у них энцефалопатии в условиях медленной вирусной инфекции (болезни Крейтц-фельдта.Якоба, куру и др.).
Другими необычными агентами, близкими к вирусам, являются вироиды – небольшие молекулы кольцевой, суперспирализованной РНК, не содержащие белка, вызывающие заболевания у растений.
Вирусы взаимодействуют с клеткой по типам:
- продуктивный тип, завершающийся образованием вирусного потомства;
- абортивный тип, не завершающийся образованием новых вирусных частиц, поскольку инфекционный процесс прерывается на одном из этапов;
- интефативный тип, или вирогения, характеризующийся встраиванием вирусной ДНК в хромосому клетки-хозяина.
Стадии репродукция вирусов:
Адсорбция.
Взаимодействие вируса с клеткой начинается с процесса адсорбции, т. е. прикрепления вирусов к поверхности клетки. Это высокоспецифический процесс. Вирус адсорбируется на определенных участках клеточной мембраны – так называемых рецепторах. Клеточные рецепторы могут иметь разную химическую природу, представляя собой белки, углеводные компоненты белков и липидов, липиды. Число специфических рецепторов на поверхности одной клетки колеблется от 104 до 105. Следовательно, на клетке могут адсорбироваться десятки и даже сотни вирусных частиц.
Способность вирусов избирательно поражать определенные клетки органов и тканей организма называют тропизмом вирусов (от греч. tropos – направление).
Проникновение в клетку.
Существует два способа проникновения вирусов животных в клетку: виропексис и слияние вирусной оболочки с клеточной мембраной. При виропексисе после адсорбции вирусов происходят инвагинация (впячивание) участка клеточной мембраны и образование внутриклеточной вакуоли, которая содержит вирусную частицу. Вакуоль с вирусом может транспортироваться в любом направлении в разные участки цитоплазмы или ядро клетки. Процесс слияния осуществляется одним из поверхностных вирусных белков капсидной или суперкапсидной оболочки. По-видимому, оба механизма проникновения вируса в клетку не исключают, а дополняют друг друга.
Биосинтез компонентов вируса. Проникшая в клетку вирусная нуклеиновая кислота несет генетическую информацию, которая успешно конкурирует с генетической информацией клетки. Она дезорганизует работу клеточных систем, подавляет собственный метаболизм клетки и заставляет ее синтезировать новые вирусные белки и нуклеиновые кислоты, идущие на построение вирусного потомства.
Реализация генетической информации вируса осуществляется в соответствии с хорошо известными из биологии процессами транскрипции (от лат.transcriptio – переписывание, т.е. синтез информационных РНК – иРНК, комплементарных матричным ДНК или РНК), трансляции (от лат. translatio – передача, т. е. синтез белков на рибосомах клетки с участием иРНК) и репликации (от лат. replicatio – повторение, т. е. синтез молекул нуклеиновой кислоты, гомологичных геному). Поскольку генетический аппарат вирусов достаточно разнообразен, то передача наследственной информации в отношении синтеза иРНК различна.
Основные схемы реализации вирусной генетической информации могут быть представлены следующим образом:
- для ДНК-содержащих вирусов: ДНК вируса -> иРНК -> белок вируса;
- для РНК-содержащих минус-нитевых вирусов: РНК вируса -> иРНК -белок вируса;
- для РНК-содержащих плюс-нитевых вирусов: РНК вируса -> белок вируса;
- для РНК-содержащих ретровирусов: РНК вируса – комплементарная ДНК -> иРНК -> белок вируса.
Для синтеза иРНК одни вирусы используют клеточные ферменты, другие – собственный набор ферментов (полимераз).
Вирусная нуклеиновая кислота кодирует синтез двух классов белков: неструктурных белков-ферментов, которые обслуживают процесс репродукции вирусов на разных его этапах, и структурных белков, которые войдут в состав вирусных частиц потомства.Синтез компонентов вируса (белков и нуклеиновых кислот) разобщен во времени и пространстве, т. е. протекает в разных структурах ядра и цитоплазмы клетки. Вот почему этот уникальный способ размножения вирусов называется дисыонктивным (от лат. disjunctus – разобщенный).
Принципы сборки вирусов, имеющих разную структуру:
- формирование вирусов является многоступенчатым процессом с образованием промежуточных форм;
- сборка просто устроенных вирусов заключается во взаимодействии молекул вирусных нуклеиновых кислот с капсидными белками и образовании нуклеокапсидов (например, вирусы полиомиелита). У сложно устроенных вирусов сначала формируются нуклеокапсиды, с которыми взаимодействуют белки суперкапсидных оболочек (например, вирусы гриппа);
- формирование вирусов происходит не во внутриклеточной жидкости, а на ядерных или цитоплазматических мембранах клетки;
- сложно организованные вирусы в процессе формирования включают в свой состав компоненты клетки-хозяина (липиды, углеводы).
Выход вирусов из клетки. Различают два основных типа выхода вирусного потомства из клетки.
Первый тип – взрывной – характеризуется одновременным выходом большого количества вирусов. При этом клетка быстро погибает. Такой способ выхода характерен для вирусов, не имеющих суперкапсидной оболочки.
Таким образом, внешняя оболочка этих вирусов формируется в процессе их выхода из клетки. При таком механизме клетка может продолжительное время продуцировать вирус, сохраняя в той или иной мере свои основные функции.Время, необходимое для осуществления полного цикла репродукции вирусов, варьирует от 5-6 ч (вирусы гриппа, натуральной оспы и др.) до нескольких суток (вирусы кори, аденовирусы и др.). Образовавшиеся вирусы способны инфицировать новые клетки и проходить в них указанный выше цикл репродукции.
Интегративный тип взаимодействия (вирогения)
Интегративный тип взаимодействия (вирогения) характеризуется встраиванием (интеграцией) нуклеиновой кислоты вируса в хромосому клетки. При этом вирусный геном реплицируется и функционирует как составная часть клеточного генома.
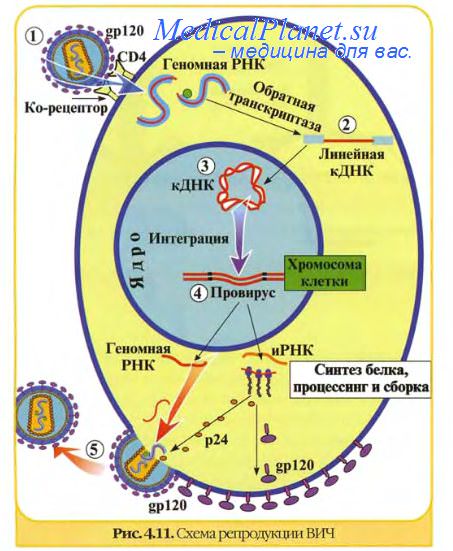
Интеграция вирусного генетического материала с ДНК клетки характерна для определенных групп вирусов: бактериофагов, опухолеродных (онкогенных) вирусов, некоторых инфекционных вирусов (вирус гепатита В, аденовирус, ВИЧ). Для интеграции с хромосомой клетки необходима кольцевая форма двунитчатой вирусной ДНК. У ДНК-содержащих вирусов (вирус гепатита В) их ДНК обладает свойством встраиваться в геном клетки при участии ряда ферментов.
У некоторых РНК-содержащих вирусов (ВИЧ, онкогенные вирусы) процесс интеграции более сложный и является обязательным в цикле их репродукции. У этих вирусов сначала на матрице РНК с помощью вирусспецифического фермента обратной транскриптазы (ревертазы) синтезируется ДНК-копия (кДНК), которая затем встраивается в ДНК клетки. ДНК вируса, находящаяся в составе хромосомы клетки, называется ДНК-провирусом. При делении клетки, сохраняющей свои нормальные функции, ДНК-провирус переходит в геном дочерних клеток, т.е. состояние вирогении наследуется. ДНК-провирус несет дополнительную генетическую информацию, в результате чего клетки приобретают ряд новых свойств.
Так, интеграция может явиться причиной возникновения ряда аутоиммунных и хронических заболеваний, разнообразных опухолей. Под воздействием ряда физических и химических факторов ДНК-провирус может исключаться из клеточной хромосомы и переходить в автономное состояние, что ведет к репродукции вируса.
Признаки медленных вирусных инфекции:
- длительный инкубационный период (в течение нескольких месяцев или лет);
- своеобразное поражение органов и тканей, преимущественно центральной нервной системы;
- медленное, но неуклонное прогрессирование заболевания;
- неизбежная смерть.
Возбудителями медленных вирусных инфекций иногда могут быть обычные вирусы (вирусы кори, краснухи и др.). Например, вирус кори может вызывать подострый склерозирующий панэнцефалит (ПСПЭ), вирус краснухи – прогрессирующую врожденную краснуху и прогрессирующий краснушный панэнцефалит. В основе патогенеза медленных вирусных инфекций лежат персистенция вируса (длительное пребывание в организме) и его повреждающее действие на клетки и ткани.
Медленные вирусные инфекции могут развиваться в результате действия не только обычных вирусов, таких, как вирусы кори, краснухи и др., но и инфекционных белковых частиц – прионов.
Читайте также:
