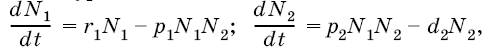Читайте также:
- А- как экономические отношения;
- Административно-правовые нормы и правоотношения
- Алексеев. Правовой статус личности — это правовое положение человека, отражающее его фактическое состояние во взаимоотношениях с обществом и государством.
- Б) сохраняют силу, кроме случаев, когда в законе установлено, что его действие распространяется на отношения, возникшие из ранее заключенных договоров;
- Билет № 4 Конституционно-правовые отношения: понятие, особенности, виды
- Билет №17. Экономико-математические модели в компьютерном управлении.
- Билет №27 Сетевые модели. Детерминированные, вероятностные модели.
- Билет №32. Критический путь в сетевой модели. Минимизация общей продолжительности проекта с учетом трудовых ресурсов.
- Блоковая модель документа. Содержимое блока, рамки, поля и отступы. Блочные и строчные теги как элементы блоковой модели. Свойства display, overflow.
- Бюджетные правоотношения и нормы бюджетного права.
|
Хищник – жертва.
Некоторые животные способны менять окраску в зависимости от цвета фона. Например, головоногие моллюски, некоторые ракообразные, рыбы, амфибии, рептилии. Экспериментально показано, что из всех рыб, выживших в водоёме, где охотился пингвин, 66% составили особи, окрашенные соответственно фону.
Взаимодействия хищников и их жертв очень динамичны и взаимообусловлены. Это особенно ярко выражается в проявлениях адаптивного поведения. Наблюдения за головастиками американской жабы Bufo americans показали, что в присутствии хищных личинок стрекоз они снижают активность, изменяется и их распределение в пространстве. Например, у коловраток Brahionus calchforus в присутствии хищной коловратки Asplanchna развиваются длинные боковые выросты панциря, которые препятствуют заглатыванию их. Образование этих шпор индуцируется веществом выделяемым хищником и уменьшением пищи.
Паразит – хозяин.
Паразитизм – это форма взаимоотношений двух различных видов организмов, носящих онтогенетический характер, при которой организмы одного вида (паразита, потребителя) живут за счёт питательных веществ или тканей организма другого вида (хозяина) в течение определённого времени. Паразитизм известен на всех уровнях организации живого, начиная с вирусов и бактерий и заканчивая высокоорганизованными животными.
Распространение паразитизма. Чем сложнее устроен организм, тем выше эффективность его гомеостазирующих систем и поэтому больше благоприятных возможностей он предоставляет в качестве среды обитания. Чем организм совершеннее, тем меньше становится для него потребность использовать благоприятные условия в другом организме. Это объясняет, почему явление паразитизма наиболее представлено среди микроорганизмов и примитивных многоклеточных. Число видов, ведущих паразитический образ жизни, резко падает у высокоорганизованных организмов. У высших животных встречается наиболее обильное разнообразие населения паразитов.
Формы паразитизма и взаимные адаптации паразитов и их хозяев.
· Эктопаразитизм (клещи, напр. таёжный клещ, собачий клещ; блохи);
· Эндопаразитизм (паразитические черви, напр. печёночный сосальщик, широкий лентец).
По степени тесноты связей паразита и хозяина выделяют 2 формы:
1. Облигатный – ведёт только паразитический образ жизни, не выживает без обязательной связи с хозяином (все черви паразиты, напр. Taeniarhynchus saginatus – бычий цепень). Характеризуется наиболее полным комплексом приспособлений к использованию организма хозяина как среды.
2. Факультативный – ведут свободный образ жизни и лишь при особых условиях переходят к паразитическому существованию. Например, Armillaria mellea – опёнок. Их связь исключительно трофическая. Морфофизиологические и биологические адаптации к паразитизму выражены слабо.
Адаптации парзитов: среда в виде живого и подвижного организма требует формирования способов фиксации в ней: зацепки, крючья, присоски, видоизменения формы тела. Эктопаразиты имеют сплющенное тело. У эндопаразитов утрачиваются органы, которые не нужны им в среде хозяина.
Математические модели.
В естественных условиях (в гетерогенной среде) возникает следующая временная и причинно-следственная цепь: размножение жертвы→размножение хищника→резкое сокращение численности жертвы→падение численности хищника→размножение жертвы и т.д.
Волны жизни хищника и жертвы следуют друг за другом с постоянным сдвигом по фазе, и в среднем численность как хищника так и жертвы остаётся примерно на одном уровне. Длительность периода зависит от скорости роста обоих видов, а так же от исходных параметров. Для популяции жертвы влияние хищника положительно, т.к. её чрезмерное размножение привело бы к краху её численности. В свою очередь, все механизмы, препятствующие полному истреблению жертвы, способствуют сохранению пищевой базы хищника.
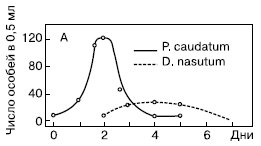
ГРАФИКИ. Взаимоотношения между хищником и жертвой: А – в гомогенной среде инфузория Dinidium nasatum погибает от голода после поедания жертвы; Б – в гетерогенной среде – жертва частично может укрыться от хищника, и он вымирает; В – естественные колебания численности хищника и жертвы ( по Гаузе).
Рис. 3. Взаимодействие между инфузорией дидиниум и инфузорией туфелькой в однородной среде (количество особей в 0,5 см3)

Рис. 5. Взаимодействие между инфузорией дидиниум и инфузорией туфелькой в неоднородной среде (количество особей в 0,5 см)

Рис. 6. Взаимодействие между инфузорией дидиниум и инфузорией туфелькой в неоднородной среде при наличии иммиграций (1 инфузория туфелька + 1 дидиниум)
Эксперименты Гаузе.
Жертва – инфузория туфелька Paramecium caudatum.
Хищник – инфузория дидиниум Didinium nasutum.
В качестве пищи для инфузории туфельки Гаузе использовал культуру сенной палочки (Bacillus subtilis).
Первая серия опытов была поставлена Гаузе в так называемой однородной среде. Для этого использовалась разведенная бактериальная культура без осадка. Особенность биологии дидиниум заключается в том, что он не является активным охотником, а просто хватает все, что попадается ему на пути. Инфузория туфелька, ускользая от хищника, использует значительно бóльшую скорость передвижения, а также особые защитные образования – трихоцисты. При этом она никогда не прячется в убежища. Совокупность этих особенностей определила чистоту данной серии опытов и позволило Гаузе выделил процесс взаимодействия хищника и жертвы в чистом виде. Бактериальная культура без осадка разливалась в пробирки объемом 0,5 мл. В нее помещалось 5 особей инфузории туфельки, а через 2 суток вносили 3 особи хищной инфузории дидиниум. Практически сразу же после внесения хищников в культуру, численность жертвы начинала убывать, поскольку дидиниум поедает новую инфузорию туфельку через каждые 3 часа. Дидиниум интенсивно размножался, достигая максимума численности к 3-м – 4-м суткам опыта. На 5-е сутки дидиниум выел всех инфузорий туфелек, а на 6-е – все хищники погибли от голода.
При первой иммиграции в свободную среду дидиниум не нашел жертвы и погиб. После этого начался интенсивный рост инфузории туфельки. К моменту второй иммиграции концентрация инфузории туфельки в среде была уже очень велика, благодаря чему начался рост популяции дидиниум. Третья иммиграция пришлась на момент интенсивного уничтожения жертв хищником, и потому не внесла существенных изменений в общую картину. К четвертой иммиграции дидиниум выел всех инфузорий туфелек, после чего погиб сам. Внесенная в среду инфузория туфелька дала начало новому циклу роста жертв. В дальнейшем, такие изменения численности обоих видов регулярно повторялись.
Проанализировав результаты, полученные в ходе трех серий опытов, Гаузе пришел к заключению, что периодические колебания численности хищника и жертвы не являются свойством процесса взаимодействия между ними, как это предполагали математики. Вероятно, они возникают в результате постоянного вмешательства извне (убежища для жертв, сохранение хищников путем инцистирования, иммиграция и эмиграция, колебания факторов среды и т. д.). Именно поэтому в естественных условиях природа периодических колебаний численности находящихся в непосредственном взаимодействии видов может быть весьма сложной.
Дата добавления: 2015-04-18 ; просмотров: 29 ; Нарушение авторских прав

УДК [619:616.995.773.4+63.001.573] : 636.32/.38
Применение математических моделей в изучении
паразитарных систем (на примере функционирования системы паразит-хозяин при эстрозе овец)
Ставропольский государственный аграрный университет
Вопрос о закономерностях динамики численности организмов имеет в экологии весьма важное значение. В прикладной энтомологии он важен в связи с тем, что дает возможность прогнозирования развития насекомых, предупреждения их массового размножения.
Система паразит-хозяин является классическим объектом математической экологии и в настоящее время.
Если же паразит использует адаптивную стратегию, то в этой системе уже может существовать устойчивое равновесие, возникают сбалансированные отношения в системе паразит-хозяин, когда обе популяции коэволюционируют.
Замечено, что в условиях неустойчивого равновесия система не разрушается, в ней появляется предельный цикл, колебания. Их возникновение существенным образом зависит от популяционных характеристик паразита. Если естественная смертность (m) паразита мала, а его плодовитость (А) велика, то равновесие почти всегда глобально устойчиво, каковы бы не были начальные значения численности паразита и паразитоносителя, система рано или поздно приходит в равновесие. Уменьшение l (l = А/m) способствует снижению устойчивости системы, появляются такие значения численности, при которых система уже не вернется к равновесию. В условиях дальнейшего уменьшения l амплитуда колебаний растет, что приводит к разрушению системы, в результате чего вымирает либо паразит, либо паразит и хозяин.
При рассмотрении более сложных моделей следует учитывать выработку защитных реакций у хозяина, групповые эффекты у паразита и др.
Система паразит-хозяин может быть устойчивой только в том случае, если в результате коэволюции хозяин становится более резистентным, а паразит менее вирулентным. Это доказано экспериментально.
Проведенный корреляционный анализ количественной зависимости трех стадий личиночного развития овечьего овода, в результате исследования позволил выявить высокую степень взаимосвязи между количеством личинок первой и второй стадий, меньшую зависимость между личинками второй и третьей стадий, независимость количественного уровня личинок третьей и первой стадий. Проведенный анализ по коэффициентам множественной регрессии показал тесную зависимость количественных показателей личинок первой стадии от численности личинок второй и третьей стадий. Наряду с методами парной корреляции, множественной регрессии был проведен анализ взаимосвязи между личинками разных стадий и методом ранговой корреляции, который относится к параметрическим методам. Такой анализ позволил подтвердить отрицательный характер корреляции между личинками первой и третьей стадий, установить очень высокую степень этой зависимости /r = -0,99/ /r 0,05). Однако, несмотря на отсутствие достоверных различий между ними, есть основание считать, что обнаруженные различия могут закономерно повторяться. В пользу этого свидетельствуют сходные закономерности в соотношении указанных средних показателей, установленные нами и другими авторами [2] при обследовании овец, инвазированных личинками полостного овода в степной зоне Северного Кавказа в период I965-I968 г. г.
Было установлено относительно высокое количество личинок 1-ой стадии у овец, достигающее по сравнению с личинками других стадий – 49%, количество личинок 2-ой стадии составляло 31%, третьей -20%. Установлены общие закономерности в кратности уменьшения личинок по мере их созревания - одинаковая кратность снижения личинок 2-ой стадии по отношению к 1-ой и 3-ей стадиям. Менее выраженная кратность уменьшения числа личинок последующих стадий, по отношению к предыдущей, обнаруженная нами (1,5 раза), по сравнению с аналогичными результатами установленными Щербань (1976) (3,5 раза), по-видимому, объясняется активно протекающим процессом развития личинок или интенсивной элиминацией последних из организма овец. Это явление может быть обусловлено неразвитым механизмом защиты у инвазированных овец. Дополнительные исследования в этой области позволят выработать, с учетом полученных данных, при выборочном подсчете у овец личинок различных стадий, показатели иммунологической реактивности поголовья овец.
Будут способствовать разработке методов оценки реактивности организма инвазированных овец, расшифровке процессов, обусловливающих количественные изменения личинок различных стадий. Рассчитанный общий парный коэффициент корреляции ( r = 0,696; р 0,05) и не зависит от количества личинок 1-ой стадии ( г =-0,131 : р >0,05).
Полученные результаты можно объяснить, обстоятельством гибели личинок 1-ой и 2-ой стадий. Количество личинок 3-ей стадии регламентировалось выходом последних в окружающую среду для окукливания. Следует отметить, что рассчитанный нами коэффициент корреляции был отрицательным, что свидетельствует об обратном характере связи.
На основе корреляционного анализа можно прогнозировать характер функционирования паразитарной системы. Однако корреляционный анализ имеет ряд недостатков. Более перспективным является регрессоионный анализ, который точнее аргументирует взаимообусловленность явлений, выявляемых при расчете парных коэффициентов корреляции. Регрессионный анализ, наряду с двусторонними связями позволяет выявить и одностороннюю зависимость между теми или иными явлениями.
Проведенный нами регрессионный анализ по коэффициентам множественной регрессии по Х (Y1/X2, X3; Y2/Х1, X3 и Y3/X1, X2), где Y1, Y2 и X1, X2, X3 соответствовал количеству личинок 1-ой, 2-ой, 3-ей стадий их развития, показал тесную зависимость количественного уровня личинок 1 стадии от численности личинок 2 и 3 стадий. Взаимосвязь между численностью личинок 1 и 3 стадий имела отрицательный характер. При сопоставлении значений парного коэффициента корреляции и регрессии, данные регрессионного анализа подтвердили наличие отрицательной связи между числом личинок указанных стадий, (коэф. детерминации r = - 0,52) выявили изменения значения показателя Y1 обусловленные изменением показателей Х1 и Х3. Степень взаимосвязей между числом личинок всех стадий достаточно высока (коэф. множественной корреляции - 0,72). При расчете коэффициентов множественной регрессии Y3 по Х1 и X2 только 23% изменений значения Y3, обусловлены изменением значений как Х1, так и X3, (коэф. детерминации - 0,23) при средней степени взаимосвязи между тремя исследуемыми показателями (коэф. множественной регрессии - 0,48), что подтверждается и более высоким коэффициентом регрессии Х3 ( 0,60), чем Х1 (0,1).
Наряду с методами парной корреляции, множественной регрессии был проведен анализ взаимосвязи между личинками различных стадий и методом ранговой корреляции, который относится к параметрическим методам, не требующих знания закона распределения единичных вариантов и используемых для относительно небольшого количества единичных определений.
Такой анализ позволил подтвердить отрицательный характер корреляции между числом личинок I и 3 стадий, установить очень высокую степень этой взаимосвязи (r = - 0,99) и достоверность (р
8.6.6. Математическое моделирование в экологии
Надорганизменные системы, которые изучает экология – популяции, биоценозы, экосистемы, – чрезвычайно сложны. В них возникает множество взаимосвязей, сила и постоянство которых непрерывно меняются. Одни и те же внешние воздействия могут привести к различным, иногда прямо противоположным результатам, в зависимости от того, в каком состоянии находилась система в момент воздействия.
Предвидеть ответные реакции системы на действие конкретных факторов можно лишь через сложный анализ существующих в ней количественных взаимоотношений и закономерностей. В экологии поэтому широкое распространение получил метод математического моделирования как средство изучения и прогнозирования природных процессов.
Суть метода заключается в том, что с помощью математических символов строится абстрактное упрощенное подобие изучаемой системы. Затем, меняя значение отдельных параметров, исследуют, как поведет себя данная искусственная система, т. е. как изменится конечный результат.
В построении математических моделей сложных процессов выделяются следующие этапы.
1. Прежде всего, те реальные явления, которые хотят смоделировать, должны быть тщательно изучены: выявлены главные компоненты и установлены законы, определяющие характер взаимодействия между ними. Если неясно, как связаны между собой реальные объекты, построение адекватной модели невозможно. На этом этапе должны быть сформулированы те вопросы, ответ на которые должна дать модель. Прежде чем строить математическую модель природного явления, надо иметь гипотезу о его течении.
2. Разрабатывается математическая теория, описывающая изучаемые процессы с необходимой детальностью. На ее основе строится модель в виде системы абстрактных взаимодействий. Установленные законы должны быть облечены в точную математическую форму. Конкретные модели могут быть представлены в аналитической форме (системой аналитических уравнений) или в виде логической схемы машинной программы. Модель природного явления есть строгое математическое выражение сформулированной гипотезы.
3. Проверка модели – расчет на основе модели и сличение результатов с действительностью. При этом проверяется правильность сформулированной гипотезы. При значительном расхождении сведений модель отвергают или совершенствуют. При согласованности результатов модели используют для прогноза, вводя в них различные исходные параметры.
Следует, однако, отметить, что сама по себе математическая модель не может служить абсолютным доказательством правильности той или иной гипотезы, так как может оказаться, что разные гипотезы приводят к сходным результатам, но она служит одним из путей анализа реальности.
Моделируют различные по характеру процессы, происходящие в реальной среде, как, например, отдельные типы экологических взаимодействий хищник – жертва, паразит – хозяин, конкурентные отношения, мутуализм и др. Математическими моделями описываются и проверяются разные варианты динамики численности, популяций, продукционные процессы в экосистемах, условия стабилизации сообществ, ход восстановления систем при разных формах нарушений и многие другие явления. Сами методы математического моделирования биологических систем развиваются, совершенствуются и разнообразятся.
Например, одну из простейших математических моделей для системы паразит – хозяин в динамике численности насекомых разработал в 1925 г. статистик А. Лотка, который вывел следующие уравнения:

где N 1– численность популяции хозяина; N 2 – численность популяции паразита; r 1– удельная скорость увеличения популяции хозяина; d 2 – удельная скорость гибели популяции паразита; p 1 и р 2– константы. График процесса паразитической инвазии, построенный по таким уравнениям, обнаруживает, что в результате взаимодействия двух видов должны возникать осцилляции (колебания) с постоянной амплитудой, которая зависит от соотношения между скоростями увеличения численности двух видов.
В период, когда были сделаны эти расчеты, экологи вели поиск причин циклических колебаний численности, которые были обнаружены к тому времени у ряда видов. Делались попытки отыскать внешние факторы (космические, солнечные, атмосферные), ответственные за периодические изменения популяций. Модели А. Лотки и В. Вольтерра позволили выдвинуть идею, что периодический колебательный режим в популяциях может возникнуть в результате межвидовых отношений и без внешнего периодического воздействия. Эта идея оказалась плодотворной для дальнейшего развития теории динамики численности популяций. Однако сама модель являлась не адекватной, т. е. не описывала действительность, так как в природе практически не обнаруживаются подобные непрерывные осцилляции с постоянной амплитудой у пар видов, связанных по типу хищник – жертва или паразит – хозяин.
В 1933 г. А. Никольсон, несколько усложнив математическую модель Лотки и введя в систему дополнительных хозяев и паразитов, показал, что это ослабляет осцилляции. В 1936 г. А. Н. Колмогоров разработал новые подходы и описал также возможности устойчивого стационарного состояния системы взаимодействующих через трофические связи видов. Позднее для систем хищник – жертва, паразит – хозяин было предложено множество других моделей. С введением в модели дополнительных параметров сильно усложняется математический аппарат и техника расчетов. Многие из этих ограничений позволило снять использование электронно-вычислительных машин.
Математическое моделирование широко применяется при решении экологических проблем, связанных с антропогенными воздействиями на природную среду. В современных математических моделях выделяют тактические и стратегические модели. Тактические модели экосистем и популяций служат для экологического прогнозирования их состояния, в том числе при разного рода экзогенных воздействиях. Стратегические модели строят в основном с исследовательскими целями, для вскрытия общих законов функционирования биологических систем, таких, как стабильность, разнообразие, устойчивость к воздействиям, способность возвращаться в исходное состояние. В задачи стратегических моделей входит изучение с помощью ЭВМ последствий разных стратегий управления экосистемами, чтобы иметь возможность выбрать оптимальную.
Модели, которые описывают взаимодействие общества и природы и в которых учитывают не только экологические, но и экономические, демографические и социальные показатели, называют эколого-экономическими моделями. Такие модели разрабатывают для долгосрочного прогнозирования экономического роста и общей оценки влияния человеческой деятельности на природную среду.
Данный текст является ознакомительным фрагментом.
Надорганизменные системы, которые изучает экология — популяции, биоценозы, экосистемы — чрезвычайно сложны. В них возникает множество взаимосвязей, сила и постоянство которых непрерывно меняются. Одни и те же внешние воздействия могут привести к различным, иногда прямо противоположным результатам, в зависимости от того, в каком состоянии находилась система в момент воздействия.
Предвидеть ответные реакции системы на действие конкретных факторов можно лишь через сложный анализ существующих в ней количественных взаимоотношений и закономерностей. В экологии поэтому широкое распространение получил метод математического моделирования как средство изучения и прогнозирования природных процессов.
Суть метода заключается в том, что с помощью математических символов строится абстрактное упрощенное подобие изучаемой системы. Затем, меняя значение отдельных параметров, исследуют, как поведет себя данная искусственная система, т. е. как изменится конечный результат.
В построении математических моделей сложных процессов выделяются следующие этапы.
1. Прежде всего, те реальные явления, которые хотят смоделировать, должны быть тщательно изучены: выявлены главные компоненты и установлены законы, определяющие характер взаимодействия между ними. Если неясно, как связаны между собой реальные объекты, построение адекватной модели невозможно. На этом этапе должны быть сформулированы те вопросы, ответ на которые должна дать модель. Прежде чем строить математическую модель природного явления, надо иметь гипотезу о его течении.
2. Разрабатывается математическая теория, описывающая изучаемые процессы с необходимой детальностью. На ее основе строится модель в виде системы абстрактных взаимодействий. Установленные законы должны быть облечены в точную математическую форму. Конкретные модели могут быть представлены в аналитической форме (системой аналитических уравнений) или в виде логической схемы машинной программы. Модель природного
явления есть строгое математическое выражение сформулированной гипотезы.
3. Проверка модели —расчет на основе модели и сличение результатов с действительностью. При этом проверяется правильность сформулированной гипотезы. При значительном расхождении сведений модель отвергают или совершенствуют. При согласованности результатов модели используют для прогноза, вводя в них различные исходные параметры.
Следует, однако, отметить, что сама по себе математическая модель не может служить абсолютным доказательством правильности той или иной гипотезы, так как может оказаться, что разные гипотезы приводят к сходным результатам, но она служит . одним из путей анализа реальности.
Моделируют различные по характеру процессы, происходящие в реальной среде, как, например, отдельные типы экологических взаимодействий хищник — жертва, паразит — хозяин, конкурентные отношения, мутуализм и др. Математическими моделями описываются и проверяются разные варианты динамики численности популяций, продукционные процессы в экосистемах, условия стабилизации сообществ, ход восстановления систем при разных формах нарушений и многие другие явления. Сами методы математического моделирования биологических систем развиваются, совершенствуются и разнообразятся.
Например, одну из простейших математических моделей для системы паразит — хозяин в динамике численности насекомых разработал в 1925″г. А. Лотки, который вывел следующие уравнения:

где Ni — численность популяции хозяина; N2 — численность популяции паразита; г — удельная скорость увеличения популяции хозяина; d2 —удельная скорость гибели популяции паразита; рхл р2 — константы. График процесса паразитической инвазии, построенный по таким уравнениям, обнаруживает, что в результате взаимодействия двух видов должны возникать осцилляции (колг-бания) с постоянной амплитудой, которая зависит от соотношения между скоростями увеличения численности двух видов.
В период, когда были сделаны эти расчеты, экологи вели поиск причин циклических колебаний численности, которые были обнаружены к тому времени у ряда видов. Делались попытки отыскать внешние факторы (космические, солнечные, атмосферные), ответственные за периодические изменения популяций. Модели А. Лотки и В. Вольтерра позволили выдвинуть идею, что периодический колебательный режим в популяциях может возникнуть в результате межвидовых отношений и без внешнего периодического воздействия. Эта идея оказалась плодотворной для дальнейшего развития теории динамики численности популяций. Однако сама модель являлась не адекватной, т. е. не описывала действительность, так как в природе практически не обнаруживаются подобные непрерывные осцилляции с постоянной амплитудой у пар видов, связанных по типу хищник — жертва или паразит — хозяин.
В 1933 г. А. Никольсон, несколько усложнив математическую модель Лотки и введя в систему дополнительных хозяев и паразитов, показал, что это ослабляет осцилляции. В 1936 г. A. >i. Кол мо гор о в разработал новые подходы и описал также возможности устойчивого стационарного состояния системы взаимодействующих через трофические связи видов. Позднее для систем хищник — жертва, паразит — хозяин было предложено множество других моделей. С введением в модели дополнительных параметров сильно усложняется математический аппарат и техника расчетов. Многие из этих ограничений позволило снять использование электронно-вычислительных машин.


С 60-х годов появляется множество работ, посвященных математическому анализу с помощью ЭВМ взаимоотношений хищник — жертва и паразит — хозяин. Экспериментирование с моделями на вычислительных машинах открыло широкие возможности для поиска стратегий управления биологическими системами. Моделирование на ЭВМ позволяет также совершенствовать систему сбора исходных сведений. Так, если модель содержит нереалистические предположения, то картина на выходе ЭВМ позволяет понять, как надо упорядочить эксперименты и наблюдения для получения необходимой количественной информации.
Модели хищник — жертва играют большую роль в планировании рыбного, китобойного, охотничьего промыслов, так как изъятие человеком части популяции диких животных с экологических позиций является аналогом природного хищничества. Предельная степень эксплуатации, которую может выдержать популяция, различна у разных видов. Важно вовремя заметить симптомы, свидетельствующие, что изъятие из популяции приближается к предельно допустимому уровню, после которого может быть нарушена ее воспроизводительная способность.
Например, по результатам машинных экспериментов со статистикой китобойного промысла в 60-х годах выявлены показатели допустимых масштабов добычи и симптомы гибельной эксплуатации популяции синих китов. Если популяция эксплуатируется интенсивно, но не чрезмерно, то в моделях обнаруживается уменьшение размеров и среднего возраста особей, кривые выживания изменяются, но не настолько, чтобы нарушалась плодовитость стада в целом. В реальной действительности были обнаружены предсказанные моделями симптомы гибельной эксплуатации китового стада — сокращение доли беременных самок, сильные изменения кривых выживания, уменьшение размеров уловов на единицу промыслового усилия, неспособность популяции быстро восстановить численность после прекращения промысла. Синих китов осталось так мало, что несмотря на международный запрет их добычи, принятый в 1967 г., поголовье остается на низком уровне и животные внесены в Красную книгу.
Моделирование трофических связей имеет большое значение для решения проблем борьбы с вредителями, регуляции численности популяций, стабилизации сообществ.
Математическое моделирование широко применяется при решении экологических проблем, связанных с антропогенными воздействиями на природную среду. В современных математических моделях выделяют тактические и стратегические модели. Тактические модели экосистем и популяций служат для экологического прогнозирования их состояния, в том числе при разного рода экзогенных воздействиях. Стратегические модели строят в основном с исследовательскими целями, для вскрытия общих законов функционирования биологических систем, таких, как стабильность, разнообразие, устойчивость к воздействиям, способность возвращаться в исходное состояние. В задачи стратегических моделей входит изучить с помощью ЭВМ последствия разных стратегий управления экосистемами, чтобы иметь возможность выбрать оптимальную.
Модели, которые описывают взаимодействие общества и природы и в которых учитывают не только экологические, но и экономические, демографические и социальные показатели, называют эколого-экономическими моделями. Такие модели разрабатывают для долгосрочного прогнозирования экономического роста и общей оценки влияния человеческой деятельности на природную среду.
Читайте также:
 Пожалуйста, не занимайтесь самолечением!При симпотмах заболевания - обратитесь к врачу.
Пожалуйста, не занимайтесь самолечением!При симпотмах заболевания - обратитесь к врачу.