
Вирусы структура взаимодействие с клеткой хозяина
Взаимодействие вируса с клеткой хозяина
Открытие вирусов Д. И. Ивановским в 1892г. положило начало развитию науки вирусологии. Более быстрому ее развитию способствовали: изобретение электронного микроскопа, разработка метода культивирования микроорганизмов в культурах клеток.
Слово “вирус” в переводе с латинского - яд (животного происхождения). Этот термин применяют для обозначения уникальных представителей живой природы, не имеющих клеточного (эукариотического или прокариотического) строения и обладающих облигатным внутриклеточным паразитизмом, т. е. которые не могут жить без клетки.
В настоящее время вирусология - бурно развивающаяся наука, что связано с рядом причин:
- ведущей ролью вирусов в инфекционной патологии человека (примеры - вирус гриппа, ВИЧ -вирус иммунодефицита человека, цитомегаловирус и другие герпесвирусы) на фоне практически полного отсутствия средств специфической химиотерапии;
- использованием вирусов для решения многих фундаментальных вопросов биологии и генетики.
Основные свойства вирусов (и плазмид), по которым они отличаются от остального живого мира.
1. Ультрамикроскопические размеры (измеряются в нанометрах). Крупные вирусы (вирус оспы) могут достигать размеров 300 нм, мелкие - от 20 до 40 нм. 1мм=1000мкм, 1мкм=1000нм.
2. Вирусы содержат нуклеиновую кислоту только одного типа - или ДНК (ДНК-вирусы) или РНК (РНК-вирусы). У всех остальных организмов геном представлен ДНК, в них содержится как ДНК, так и РНК.
3. Вирусы не способны к росту и бинарному делению.
4. Вирусы размножаются путем воспроизводства себя в инфицированной клетке хозяина за счет собственной геномной нуклеиновой кислоты.
5. У вирусов нет собственных систем мобилизации энергии и белок - синтензирующих систем, в связи с чем вирусы являются абсолютными внутриклеточными паразитами.
6. Средой обитания вирусов являются живые клетки - бактерии (это вирусы бактерий или бактериофаги), клетки растений, животных и человека.
Все вирусы существуют в двух качественно разных формах: внеклеточной - вирион и внутриклеточной - вирус. Таксономия этих представителей микромира основана на характеристике вирионов - конечной фазы развития вирусов.
Строение (морфология) вирусов
1. Геном вирусов образуют нуклеиновые кислоты, представленные одноцепочечными молекулами РНК (у большинства РНК -вирусов) или двухцепочечными молекулами ДНК (у большинства ДНК -вирусов).
2. Капсид - белковая оболочка, в которую упакована геномная нуклеиновая кислота. Капсид состоит из идентичных белковых субъединиц - капсомеров. Существуют два способа упаковки капсомеров в капсид - спиральный (спиральные вирусы) и кубический (сферические вирусы).
При спиральной симметрии белковые субъединицы располагаются по спирали, а между ними, также по спирали, уложена геномная нуклеиновая кислота (нитевидные вирусы). При кубическом типе симметрии вирионы могут быть в виде многогранников, чаще всего - двадцатигранники - икосаэдры.
3. Просто устроенные вирусы имеют только нуклеокапсид, т. е. комплекс генома с капсидом и называются “голыми”.
4. У других вирусов поверх капсида есть дополнительная мембраноподобная оболочка, приобретаемая вирусом в момент выхода из клетки хозяина - суперкапсид. Такие вирусы называют “одетыми”.
Кроме вирусов, имеются еще более просто устроенные формы способных передаваться агентов - плазмиды, вироиды и прионы.
Основные этапы взаимодействия вируса с клеткой хозяина
1. Адсорбция - пусковой механизм, связанный со взаимодействием специфических рецепторов вируса и хозяина (у вируса гриппа - гемагглютинин, у вируса иммунодефицита человека - гликопротеин gp 120).
2. Проникновение - путем слияния суперкапсида с мембраной клетки или путем эндоцитоза (пиноцитоза).
3. Освобождение нуклеиновых кислот - “раздевание” нуклеокапсида и активация нуклеиновой кислоты.
4. Синтез нуклеиновых кислот и вирусных белков, т. е. подчинение систем клетки хозяина и их работа на воспроизводство вируса.
5. Сборка вирионов - ассоциация реплицированных копий вирусной нуклеиновой кислоты с капсидным белком.
6. Выход вирусных частиц из клетки, приобретения суперкапсида оболочечными вирусами.
Исходы взаимодействия вирусов с клеткой хозяина
1. Абортивный процесс - когда клетки освобождаются от вируса:
- при инфицировании дефектным вирусом, для репликации которого нужен вирус - помощник, самостоятельная репликация этих вирусов невозможна (так называемые вирусоиды). Например, вирус дельта (D) гепатита может реплицироваться только при наличии вируса гепатита B, его Hbs - антигена, аденоассоциированный вирус - в присутствии аденовируса);
- при инфицировании вирусом генетически нечувствительных к нему клеток;
- при заражении чувствительных клеток вирусом в неразрешающих условиях.
2. Продуктивный процесс - репликация (продукция) вирусов:
- гибель (лизис) клеток (цитопатический эффект) - результат интенсивного размножения и формирования большого количества вирусных частиц - характерный результат продуктивного процесса, вызванного вирусами с высокой цитопатогенностью. Цитопатический эффект действия на клеточные культуры для многих вирусов носит достаточно узнаваемый специфический характер;
- стабильное взаимодействие, не приводящее к гибели клетки (персистирующие и латентные инфекции) - так называемая вирусная трансформация клетки.
3. Интегративный процесс - интеграция вирусного генома с геномом клетки хозяина. Это особый вариант продуктивного процесса по типу стабильного взаимодействия. Вирус реплицируется вместе с геномом клетки хозяина и может длительно находиться в латентном состоянии. Встраиваться в ДНК - геном хозяина могут только ДНК - вирусы (принцип “ДНК - в ДНК”). Единственные РНК - вирусы, способные интегрироваться в геном клетки хозяина - ретровирусы, имеют для этого специальный механизм. Особенность их репродукции - синтез ДНК провируса на основе геномной РНК с помощью фермента обратной транскриптазы с последующим встраиванием ДНК в геном хозяина.
Основные методы культивирования вирусов
1. В организме лабораторных животных.
2. В куриных эмбрионах.
3. В клеточных культурах - основной метод.
Типы клеточных культур
1. Первичные (трипсинизированные) культуры - фибробласты эмбриона курицы (ФЭК), человека (ФЭЧ), клетки почки различных животных и т. д. Первичные культуры получают из клеток различных тканей чаще путем их размельчения и трипсинизации, используют однократно, т. е. постоянно необходимо иметь соответствующие органы или ткани.
2. Линии диплоидных клеток пригодны к повторному диспергированию и росту, как правило не более 20 пассажей (теряют исходные свойства).
3. Перевиваемые линии (гетероплоидные культуры), способны к многократному диспергированию и перевиванию, т. е. к многократным пассажам, наиболее удобны в вирусологической работе - например, линии опухолевых клеток Hela, Hep и др.
Специальные питательные среды для культур клеток
Используются разнообразные синтетические вирусологические питательные среды сложного состава, включающие большой набор различных факторов роста - среда 199, Игла, раствор Хэнкса, гидролизат лактальбумина. В среды добавляют стабилизаторы рН (Hepes), различные в видовом отношении сыворотки крови (наиболее эффективной считают эмбриональную телячью сыворотку), L -цистеин и L -глютамин.
В зависимости от функционального использования среды могут быть ростовые (с большим содержанием сыворотки крови) - их используют для выращивания клеточных культур до внесения вирусных проб, и поддерживающие (с меньшим содержанием сыворотки или ее отсутствием) - для содержания инфицированных вирусом клеточных культур.
Выявляемые проявления вирусной инфекции клеточных культур
1. Цитопатический эффект.
2. Выявление телец включений.
3. Выявление вирусов методом флюоресцирующих антител (МФА), электронной микроскопией, авторадиографией.
4. Цветная проба. Обычный цвет используемых культуральных сред, содержащих в качестве индикатора рН феноловый красный, при оптимальных для клеток условиях культивирования (рН около 7,2) - красный. Размножение клеток меняет рН и соответственно - цвет среды с красного на желтый за счет смещения рН в кислую сторону. При размножении в клеточных культурах вирусов происходит лизис клеток, изменения рН и цвета среды не происходит.
5. Выявление гемагглютинина вирусов - гемадсорбция, гемагглютинация.
6. Метод бляшек (бляшкообразования). В результате цитолитического действия многих вирусов на клеточные культуры образуются зоны массовой гибели клеток. Выявляют бляшки - вирусные “ клеточно - негативные” колонии.
Название семейства вирусов заканчивается на “viridae”, рода - “virus”, для вида обычно используют специальные названия, например - вирус краснухи, вирус иммунодефицита человека - ВИЧ, вирус парагриппа человека типа 1 и т. д.
Вирусы бактерий (бактериофаги)
Естественной средой обитания фагов является бактериальная клетка, поэтому фаги распространены повсеместно (например, в сточных водах). Фагам присущи биологические особенности, свойственные и другим вирусам.
Наиболее морфологически распространенный тип фагов характеризуется наличием головки - икосаэдра, отростка (хвоста) со спиральной симметрией (часто имеет полый стержень и сократительный чехол), шипов и отростков (нитей), т. е. внешне несколько напоминают сперматозоид.
Взаимодействие фагов с клеткой (бактерией) строго специфично, т. е. бактериофаги способны инфицировать только определенные виды и фаготипы бактерий.
Основные этапы взаимодействия фагов и бактерий
1. Адсорбция (взаимодействие специфических рецепторов).
2. Внедрение вирусной ДНК (инъекция фага) осуществляется за счет лизирования веществами типа лизоцима участка клеточной стенки, сокращения чехла, вталкивания стержня хвоста через цитоплазматическую мембрану в клетку, впрыскивание ДНК в цитоплазму.
3. Репродукция фага.
4. Выход дочерних популяций.
Основные свойства фагов
Различают вирулентные фаги, способные вызвать продуктивную форму процесса, и умеренные фаги, вызывающие редуктивную фаговую инфекцию (редукцию фага). В последнем случае геном фага в клетке не не реплицируется, а внедряется (интегрируется) в хромосому клетки хозяина (ДНК в ДНК), фаг превращается в профаг. Этот процесс получил название лизогении. Если в результате внедрения фага в хромосому бактериальной клетки она приобретает новые наследуемые признаки, такую форму изменчивости бактерий называют лизогенной (фаговой) конверсией. Бактериальную клетку, несущую в своем геноме профаг, называют лизогенной, поскольку профаг при нарушении синтеза особого белка - репрессора может перейти в литический цикл развития, вызвать продуктивную инфекцию с лизисом бактерии.
Умеренные фаги имеют важное значение в обмене генетическим материалом между бактериями - в трансдукции (одна из форм генетического обмена). Например, способностью вырабатывать экзотоксин обладают только возбудитель дифтерии, в хромосому которого интегрирован умеренный профаг, несущий оперон tox, отвечающий за синтез дифтерийного экзотоксина. Умеренный фаг tox вызывает лизогенную конверсию нетоксигенной дифтерийной палочки в токсигенную.
По спектру действия на бактерии фаги разделяют на:
- поливалентные (лизируют близкородственные бактерии, например сальмонеллы);
- моновалентные (лизируют бактерии одного вида);
- типоспецифические (лизируют только определенные фаговары возбудителя).
На плотных средах фаги обнаруживают чаще с помощью спот (spot) - теста (образование негативного пятна при росте колоний) или методом агаровых слоев (титрования по Грациа).
Практическое использование бактериофагов.
1. Для идентификации (определение фаготипа).
2. Для фагопрофилактики (купирование вспышек).
3. Для фаготерапии (лечение дисбактериозов).
4. Для оценки санитарного состояния окружающей среды и эпидемиологического анализа.
Взаимодействие вируса с клеткой хозяина - это сложный многоступенчатый процесс, который начинается с адсорбции вирусных частиц на рецепторах клетки хозяина и продолжается после их проникновения внутрь клетки. В результате такого взаимодействия развивается либо продуктивная, либо абортивная, либо интегративная форма клеточной инфекции.
При продуктивной форме происходит размножение, точнее репродукция вируса, при абортивной - ее нарушение на одном из этапов, при интегративной - интеграция вирусной нуклеиновой кислоты в клеточный геном.
Вирусы - самореплицирующая форма, неспособная к бинарному делению, в отличие от микроорганизмов с клеточной организацией.
Размножение, или репродукция вирусов происходит путем репликации их нуклеиновой кислоты, и биосинтеза белков с последующей самосборкой вириона.
Вирусная репродукция представляет собой уникальную форму выражения чужеродной (вирусной) информации в клетках человека и животных, насекомых, растений и бактерий, которая состоит в подчинении клеточных матрично-генетических механизмов вирусной информации.
Жизненный цикл вирусов начинается с их адсорбции на мембране клетки -мишени и заканчивается выходом вновь синтезированных вирионов из клетки. Цикл включает в себя следующие стадии:
2. Внедрение вириона в клетку, сопряженное с одновременным разрушением его суперкапсидных и капсидных белков и высвобождением его геномной нуклеиновой кислоты.
3. Внутриклеточное размножение вируса, включающее в себя целую серию последовательных событий, заканчивающихся формированием зрелых вирионов и выходом их из клетки.
Адсорбциявируса на мембране клетки является пусковым моментом реализации его патогенных свойств, ибо без этого вирус не может проникнуть в клетку и размножаться в ней, он просто обречен на гибель.
Для каждого вируса на мембране клеток существуют специфические рецепторы, с которыми вирусы связываются с помощью своих рецепторов на основе так называемой органотропности вирусов. В связи с разнообразием клеточных и вирусных рецепторов на одних и тех же клетках могут адсорбироваться разные вирусы. Например, рецепторами для вируса гриппа являются мукопептиды, содержащие свободную N -ацетилнейраминовую кислоту, а рецептором вируса, распознающим его, является белок - гемагглютинин.
Проникновение вируса в клетку.Известны два механизма проникновения вируса в клетку:
- посредством слияния суперкапсида вируса с мембраной клетки. Благодаря этому происходит высвобождение нуклеокапсида в цитоплазму с последующей реализацией свойств вирусного генома.
- механизм пиноцитоза (виропексиса). В этом случае вирус связывается со специфическими рецепторами в области "окаймленной" ямки. Она впячивается внутрь клетки и превращается в окаймленный пузырек. Пузырек, содержащий поглощенный вирион, быстро сливается с промежуточным пузырьком - эндосомой, который сливается с лизосомой. Благодаря особым свойствам вирусных суперкапсидных белков происходит слияние липидных слоев суперкапсида и мембраны лизосомы, в результате этого нуклеокапсид оказывается в цитолизе клетки, где происходит дальнейшее "раздевание" нуклеокапсида и высвобождение геномной нуклеиновой кислоты.
Внутриклеточное размножение.Проникнув в клетку, вирусный геном полностью подчиняет жизнь клетки своим интересам и с помощью ее белоксинтезирующей системы и систем генераций энергии осуществляет собственное воспроизводство, очень часто ценой жизни клетки.
Транскрипция и репликация вирусных геномов осуществляется несколькими путями:
- Вирусы с негативным геномом РНК (минус-нитевые) имеют в своем составе вирусспецифическую РНК-полимеразу или транскриптазу. Они синтезируют и РНК на матрице геномной РНК. Подобный фермент отсутствует в нормальных клетках, но синтезируется клетками, зараженными вирусами.
- У вирусов с положительным геномом РНК (плюс-нитевые) функцию и РНК выполняет сам геном, который транслирует содержащуюся в нем информацию на рибосомы хозяиа.
- РНК - содержащие ретровирусы стоят особняком, так как в их составе есть фермент обратная транскриптаза или ревертаза, способная переписыать информацию с РНК на ДНК.
- Репликация вирусной ДНК происходит на обеих нитях ДНК при участии ДНК - полимеразы.
Сборка вириона состоит прежде всего в образовании нуклеокапсидов. Поскольку синтез вирусных белков и нуклеиновых кислот происходит в разных структурах клетки. Необходима транспортировка составных частей вириона в одно место сборки. При этом вирусные белки и нуклеиновые кислоты обладают способностью узнавать и самопроизвольно соединяться друг с другом. Многие простые вирионы собираются на репликативных комплексах - мембранах цитоплазматического ретикулума. У сложных вирионов сборка нуклеокапсида начинается на репликативных комплексах, а заканчивается на плазматической мембране. После отделения почки, содержащей нуклеокапсид и суперкапсидные белки, образуются свободные вирионы. Они через клеточную плазматическую мембрану проходят во внеклеточное пространство, дальнейшее формирование вириона происходит в мембранах цитоплазматического ретикулума и в аппарате Гольджи, откуда вирус транспортируется на поверхность клетки.
Выход вирусных частициз клетки - происходит двумя путями - простые вирусы, лишенные суперкапсида, например, пикорнавирусы вызывают деструкцию клетки и попадают во внеклеточное пространство.
Другие вирусы, имеющие липопротеидную оболочку, выходят из клетки путем почкования, в результате чего она еще сохраняет свою жизнеспособность (вирус гриппа и др.).
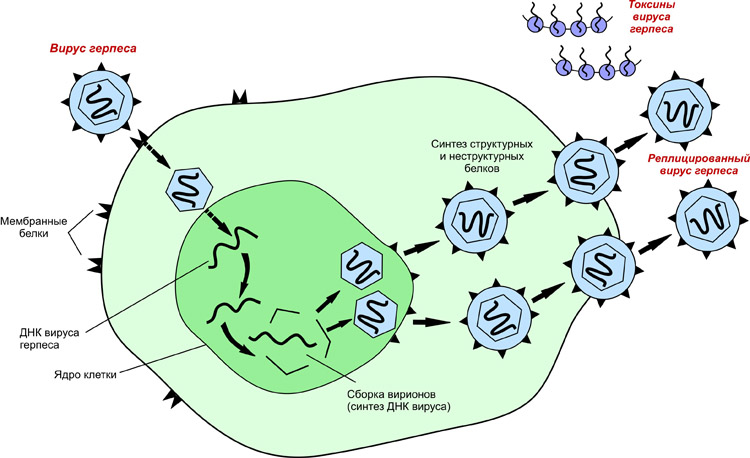
Этапы взаимодействия вируса с клеткой хозяина:
1. Адсорбция – пусковой механизм, связанный с прикреплением вириона к поверхности клетки. Выделяют две основных фазы адсорбции:
а) неспецифическая обусловлена ионным притяжением между вирусом и клеткой.
б) высокоспецифическая обусловлена гомологией, комплементарностью специфических рецепторов вируса и хозяина (у вируса гриппа – гемагглютинин к сиаловой кислоте гликопротеинов клеток дыхательных путей, у вируса иммунодефицита человека - гликопротеин gp 120 - к СД4 рецепторам Т-хелперов, моноцитов, макрофагов).
Проникновение
а) путем слияния оболочки вириона с мембраной клетки, характерно для некоторых оболочечных вирусов (парамиксовирусов, ретровирусов, герпесвирусов)
б) путем эндоцитоза (пиноцитоза) в результате захватывания и поглощения вириона клеткой: клеточная мембрана с прикрепленным вирионом впячивается с образованием внутриклеточной вакуоли (эндосомы), содержащей вирус.
3. Освобождение нуклеиновых кислот (депротеинизация) – “раздевание” нуклеокапсида и активация нуклеиновой кислоты. В результате депротеинизации удаляются поверхностные структуры вируса и высвобождается его внутренний компонент, способный вызвать инфекционный процесс.
4. Синтез нуклеиновых кислот и вирусных белков, т.е. подчинение систем клетки хозяина и их работа на воспроизводство вируса. Синтез нуклеиновых кислот и белков вируса разобщен во времени и пространстве, т.к. осуществляется в разных частях клетки. Такой способ размножения вирусов называется дизъюнктивным.
Синтез белков в клетке осуществляется благодаря процессам транскрипции – переписывания генетической информации с нуклеиновой кислоты в нуклеотидную последовательность информационной РНК (иРНК или мРНК) и трансляции – механизмов, при помощи которых последовательность нуклеотидных оснований мРНК переводится в специфическую последовательность аминокислот в синтезируемом полипептиде на рибосомах клетки хозяина.
5. Сборка вирионов – многоступенчатый процесс, включающий в себя соединение всех компонентов вириона.
Сборка просто устроенных вирусов заключается во взаимодействии вирусных нуклеиновых кислот с капсидными белками и в образовании нуклеокапсидов. У сложно устроенных вирусов сначала формируются нуклеокапсиды, которые взаимодействуют с модифицированными мембранами клеток (будущей липопротеиновой оболочкой вируса). У минус-нитевых РНК вирусов в сборку вовлекается матриксный белок (М-белок), который расположен под модифицированной клеточной мембраной.
6. Выход вирусных частиц из клетки взрывным путем или почкованием, экзоцитозом. Полный цикл репродукции вирусов завершается через 5-6 ч (вирус гриппа) или через несколько суток (вирус кори). По взрывному пути выходят из клетки просто устроенные вирусы, не имеющие липопротеиновой оболочки. Из погибающей клетки одновременно выходит большое количество вирионов. Почкование, экзоцитоз характерен вирусам, имеющим липопротеиновую оболочку, которая является производной клеточных мембран. Сначала образовавшийся нуклеокапсид транспортируется к клеточным мембранам, в которые уже встроены вирусоспецифические белки. Затем в области контакта нуклеокапсида с клеточной мембраной начинается выпячивание этих участков. Сформировавшаяся почка отделяется от клетки в виде сложно устроенного вируса. Клетка способна длительное время сохранять жизнеспособность и продуцировать вирусное потомство.
Исходы взаимодействия вирусов с клеткой хозяина
1. Продуктивный процесс - репликация (продукция) вирусов. Выделяют:
· продуктивный цитолитический процесс с образованием инфекционного потомства и выходом вирионов во внеклеточную среду.
· продуктивной нецитолитический процесс с образованием инфекционных вирусных частиц без лизиса клетки, которая продолжает функционировать.
Цитопатический эффект является результатом интенсивного размножения и формирования большого количества вирусных частиц (характерный результат продуктивного процесса), вызванного вирусами с высокой цитопатогенностью. Цитопатический эффект действия на клеточные культуры для многих вирусов носит узнаваемый специфический характер;
2. Интегративный процесс - интеграция вирусного генома с геномом клетки хозяина (вирогения). Это особый вариант продуктивного процесса по типу стабильного взаимодействия. Вирус реплицируется вместе с геномом клетки хозяина и может длительно находиться в латентном состоянии. Встраиваться в ДНК - геном хозяина могут только ДНК- вирусы (принцип “ДНК - в ДНК”). Единственные РНК - вирусы, способные интегрироваться в геном клетки хозяина - ретровирусы, имеют для этого специальный механизм. Синтез ДНК провируса на основе геномной РНК с помощью фермента обратной транскриптазы (ревертазы) с последующим встраиванием ДНК в геном хозяина.
3. Абортивный процесс - клетки освобождаются от вируса, не вызывая появление дочерней популяции или появление в меньшем количестве. При инфицировании дефектным вирусом, для репликации которого нужен вирус- помощник, самостоятельная репликация этих вирусов невозможна. Например, дельта вирус или вирус гепатита D может реплицироваться в клетке только при наличии вируса гепатита B, его HВs – антигена. При инфицировании вирусом генетически нечувствительных к нему клеток или при заражении чувствительных клеток вирусом в неблагоприятных условиях.
Возможные последствия инфекционного процесса, вызванного вирусами для клетки:
- сохранение жизнеспособности клетки
- деструкция клетки, возникающая при цитолитической инфекции (цитопатогенное действие вируса - ЦПД)
- образование многоядерных клеток в результате их слияния (симпластообразование)
- образование вирусных внутриклеточных включений
· онкогенная трансформация клетки при интеграции вирусного генома с геномом клетки (вирогении, интегративной инфекции)
Дата добавления: 2019-09-13 ; просмотров: 583 ;
Продуктивная инфекция. Репродукция вирусов
Как отмечалось выше, вирусы являются самореплицирующейся формой, неспособной к бинарному делению, в отличие от микроорганизмов с клеточной организацией. В 50-х годах было установлено, что размножение, или репродукция, вирусов происходит путем репликации их нуклеиновой кислоты и биосинтеза белков с последующей самосборкой вириона. Этот процесс происходит в разных частях клетки - ядре или цитоплазме, вследствие чего получил название дизъюнктивного, т.е. разобщенного размножения.
Вирусная репродукция представляет собой уникальную форму выражения чужеродной (вирусной) информации в клетках человека и животных, насекомых, растений и бактерий, которая состоит в подчинении клеточных матрично-генетических механизмов вирусной информации.
1-я стадия - адсорбция - характеризуется прикреплением вириона к клеточным рецепторам, представляющим собой гликопротеины клеточной мембраны, содержащей нейраминовую кислоту. Такие рецепторы имеются у ряда клеток, в частности эритроцитов, на которых адсорбируются многие вирусы. Для орто- и парамиксовирусов специфическими рецепторами являются гликолипиды, содержащие сиаловую кислоту (ганглиозиды), для других - белки или липиды клеточной мембраны. Рецепторами вирусов являются так называемые прикрепительные белки, располагающиеся в составе капсидов простых вирионов и суперкапсидов сложных вирионов. Они могут иметь форму нитей (фибры у аденовирусов) или шипов (гликопротеиновые образования на внешней оболочке орто- и парамиксо-, рабдо-, арено- и буньявирусов). Первый этап адсорбции определяется неспецифическими силами межмолекулярного притяжения, второй - специфической структурной гомологией или комплементарностью рецепторов чувствительных клеток и вирусов.
2-я стадия - проникновение вируса в клетку хозяина - происходит несколькими путями.
Рецепторно-опосредованный эндоцитоз характеризуется образованием в месте взаимодействия вириона с клеточным рецептором окаймленных пузырьков, в формировании которых принимают участие белки-клатрины.
Виропексис. Этим путем в клетку проникают сложноустроен-ные вирусы. Он заключается в слиянии мембран - вирусного суперкапсида с клеточной или ядерной мембраной. Данный процесс происходит при помощи специального бека слияния - F-белка, который находится в суперкапсиде. В результате виропексиса капсид оказывается в клетке хозяина, а суперкапсид вместе с белком встраивается в ее плазматическую мембрану (вследствие чего клетка приобретает способность сливаться с другими клетками, что приводит к передаче вируса этим клеткам).
Фагоцитоз . Данным путем вирус проникает в фагоцитирующие клетки, что приводит к незавершенному фагоцитозу.
3-я стадия - транспорт вируса внутри клетки. Он происходит с помощью внутриклеточных мембранных пузырьков, в которых вирус переносится на рибосомы, эндоплазматическую сеть или в ядро.
5-я стадия называется эклипс-фазой, которая характеризуется исчезновением вириона, поскольку он перестает обнаруживаться при элданой микроскопии. В эту стадию начинается синтез компонентов вириона, т.е. его репродукция. Она носит дизъюнктивный (раздельный) характер, поскольку компоненты вириона синтезируются в разных частях клетки: белки на рибосомах, нуклеиновые кислоты в ядре или цитоплазме. Вирус использует для этого генетический аппарат клетки, подавляя необходимые ей самой синтетические реакции.
Эта стадия начинается с транскрипции и репликации вирусного генома. Транскрипция вирусного генома двунитевых ДНК-содержащих вирусов происходит, так же как и клеточного генома, по триаде ДНК- иРНК- белок. Различия касаются только происхождения фермента ДНК-зависимой РНК-полимеразы, необходимой для данного процесса. У вирусов, геном которых транскрибируется в цитоплазме клетки хозяина (например, вирус оспы), имеется собственная вирусспецифическая РНК-полимераза. Вирусы, геномы которых транскрибируются в ядре (папова- и аденовирусы, вирусы герпеса), используют содержащуюся там клеточную РНК-полимеразу II или III.
У РНК-содержащих вирусов транскрипция их генома осуществляется несколькими путями:
1. Вирусы с негативным геномом (минус-нитевые), к которым относятся орто-, парамиксо- и рабдовирусы, имеют в своем составе вирусспецифическую РНК-полимеразу или транскриптазу. Они синтезируют иРНК на матрице геномной РНК. Подобный фермент отсутствует в нормальных клетках, но синтезируется клетками, зараженными вирусами. Он находится в составе как однонитевых, так и двунитевых РНК-содержащих вирусов.
2. У вирусов с положительным геномом (плюс-нитевые), к которым относятся пикорна-, тогавирусы и др., функцию иРНК выполняет сам геном, который транслирует содержащуюся в нем информацию на рибосомы клетки хозяина.

Вирусы — это микроскопические патогены, заражающие клетки живых организмов для самовоспроизводства. Они состоят из одного вида нуклеиновой кислоты (или ДНК или РНК, но не обе вместе), которая защищена оболочкой, содержащей белки, липиды, углеводы или их комбинацию. Размер типичного вируса варьируется от 15 до 350 нм, поэтому его можно увидеть только с помощью электронного микроскопа.
В 1892 году русский ученый Д.И. Ивановский впервые доказал существование ранее неизвестного типа возбудителя болезней, это был вирус мозаичной болезни табака. А в 1898 году Фридрих Лоффлер и Пол Фрош нашли доказательства того, что причиной ящура у домашнего скота была инфекционная частица, которая меньше, чем любая бактерия. Это были первые шаги к изучению природы вирусов, генетических образований, которые лежат где-то в серой зоне между живыми и неживыми состояниями материи. На текущий момент описано около 6 тыс. вирусов, но их существует несколько миллионов.
Строение вирусов
Вне клеток-хозяев вирусы существуют в виде белковой оболочки (капсида), иногда заключенного в белково-липидную мембрану. Капсид обволакивает собой либо ДНК, либо РНК, которая кодирует элементы вируса. Находясь в такой форме вне клетки, вирус метаболически инертен и называется вирионом.
Простая структура, отсутствие органелл и собственного метаболизма позволяет некоторым вирусам кристаллизоваться, т.е. они могут вести себя подобно химическим веществам. С появлением электронных микроскопов было установлено, что их кристаллы состоят из тесно прижатых друг к другу нескольких сотен миллиардов частиц. В одном кристалле вируса полиомиелита столько частиц, что ими можно заразить не по одному разу всех жителей Земли.
Формы вирусов
Вирусы встречаются в трех основных формах. Они бывают:
- Сферическими (кубическими или полигидральными). Вирусы герпеса, типулы, полиомы и т.д.
- Спиральными (цилиндрическими или стержнеобразными). Вирусы табачной мозаики, гриппа, эпидемического паротита и др.
- Сложными. Например, бактериофаги.
Проникновение вирусов в клетку-хозяина
Капсид в основном защищает нуклеиновую кислоту от действия клеточного нуклеазного фермента. Но некоторые белки капсида способствуют связыванию вируса с поверхностью клеток-хозяев, и работают, как ключики, вставляемые в нужные замочки. Другие поверхностные белки действуют как ферменты, они растворяют поверхностный слой клетки-хозяина и таким образом помогают проникновению нуклеиновой кислоты вируса в клетку-хозяина.
Жизненный цикл вирусов сильно отличается у разных видов, но существует шесть основных этапов жизненного цикла вирусов:
Присоединение к клетке-хозяину представляет собой специфическое связывание между вирусными капсидными белками и рецепторами на клеточной поверхности. Эта специфика определяет хозяина вируса.
Проникновение следует за прикреплением: вирионы проникают в клетку-хозяина через рецептор-опосредованный эндоцитоз или слияние мембран. Это часто называют вирусной записью.
Проникновение вирусов в клетку достигается за счет:
Размножение вирусов
После того, как вирусный геном освобождается от капсида, начинается его транскрипция или трансляция. Именно эта стадия вирусной репликации сильно различается между ДНК- и РНК-вирусами и вирусами с противоположной полярностью нуклеиновой кислоты. Этот процесс завершается синтезом новых вирусных белков и генома (точных копий внедрённых).
Механизм репликации зависит от вирусного генома.
- ДНК-вирусы обычно используют белки и ферменты клетки-хозяина для получения дополнительной ДНК, она транскрибируется в РНК-мессенджер (мРНК), которая затем используется для управления синтезом белка.
- РНК-вирусы обычно используют ядро РНК в качестве матрицы для синтеза вирусной геномной РНК и мРНК. Вирусная мРНК направляет клетку-хозяина на синтез вирусных ферментов и капсидных белков и сборку новых вирионов. Конечно, есть исключения из этого шаблона. Если клетка-хозяин не обеспечивает ферменты, необходимые для репликации вируса, вирусные гены предоставляют информацию для прямого синтеза отсутствующих белков.
Чтобы преобразовать РНК в ДНК, вирусы должны содержать гены, которые кодируют вирус-специфический фермент обратной транскриптазы. Она транскрибирует матрицу РНК в ДНК. Обратная транскрипция никогда не происходит в неинфицированных клетках. Необходимый фермент, обратная транскриптаза, происходит только от экспрессии вирусных генов в инфицированных клетках.
Вироиды
Вироиды заражают только растения. Одни вызывают экономически важные заболевания сельскохозяйственных культур, в то время как другие являются доброкачественными. Двумя примерами экономически важных вироидов являются кокосный cadang-cadang (он вызывает массовую гибель кокосовых пальм) и вироид рубцовой кожицы яблок, который безнадежно портит товарный вид яблок.
30 известных вироидов были классифицированы в две семьи.
- Члены семейства Pospiviroidae, названные по имени вироида клубневого веретена картофеля, имеют палочковидную вторичную структуру с небольшими одноцепочечными областями, имеет центральную консервативную область, и реплицируются в ядре клетки.
- Avsunviroidae, названный в честь вироида авокадо, имеет как палочковидную, так и разветвленную области, но не имеет центральной консервативной области и реплицируется в хлоропластах растительной клетки.
В отличие от вирусов, которые являются паразитами механизма трансляции хозяина, вироиды являются паразитами клеточных транскрипционных белков.
Бактериофаги

Существуют тысячи разновидностей фагов, каждый из которых может заразить только один тип или несколько близких типов бактерий или архей. Фаги классифицируются по ряду семейств вирусов; например:
Как и все вирусы, фаги являются простыми организмами, которые состоят из ядра генетического материала (нуклеиновой кислоты), окруженного капсидом белка. Нуклеиновая кислота может представлять собой либо ДНК, либо РНК, и может быть двухцепочечной или одноцепочечной.
Существует три основных структурных формы фага:
- Икосаэдрическая (20-сторонняя) головка с хвостом
- Икосаэдрическая головка без хвоста
- Нитевидная форма
Во время заражения фаг прикрепляется к бактерии и вставляет в нее свой генетический материал. После этого фаг обычно следует одному из двух жизненных циклов: литическому (вирулентному) или лизогенному (умеренному).
Литические, или вирулентные, фаги захватывают механизм клетки, чтобы скопировать компоненты фага. Затем они разрушают или лизируют клетку, высвобождая новые частицы фага.
Лизогенные, или умеренные, фаги включают свою нуклеиновую кислоту в хромосому клетки-хозяина и реплицируются с ней как единое целое, не разрушая клетку. При определенных условиях лизогенные фаги могут индуцироваться в соответствии с литическим циклом.
Существуют и другие жизненные циклы, в т.ч. псевдолизогенез и хроническая инфекция. При псевдолизогении бактериофаг проникает в клетку, но не использует механизм репликации клеток и не интегрируется в геном хозяина, просто как бы прячется внутри бактерии, не нанося ей никакого вреда. Псевдолизогенез возникает, когда клетка-хозяин сталкивается с неблагоприятными условиями роста и, по-видимому, играет важную роль в выживании фага, обеспечивая сохранение генома фага до тех пор, пока условия роста хозяина снова не станут благоприятными.
При хронической инфекции новые фаговые частицы образуются непрерывно и длительно, но без явного уничтожения клеток.
Вскоре после открытия фаги начали использовать для лечения бактериальных заболеваний человека, таких как бубонная чума и холера. Но фаговая терапия тогда не была успешной, и после открытия антибиотиков в 1940-х годах она была практически заброшена. Однако с появлением устойчивых к антибиотикам бактерий терапевтическому потенциалу фагов уделяется все больше внимания.
Наше время с антибиотиками заканчивается. В 2016 году женщина в штате Невада умерла от бактериальной инфекции, вызванной Klebsiella pneumoniae, которая была устойчивой ко всем известным антибиотикам. Бактерии, устойчивые к колистину, антибиотику последней инстанции, были обнаружены на свинофермах в Китае. В настоящее время бактерии приспосабливаются к антибиотикам быстрее, чем когда-либо.
Покажите ножницы которыми вирусы разрезают молекулу РНК что бы встроиться для мутации.Может что нибудь придумаете другое.К примеру деление цепочка аминокислот получив энергию из вне как одноименные заряды распадается на две. К каждой соединятся только те какие были ранее (другие проскочат мимо),казалось бы копии,но внутренняя энергия разная(уменьшается увеличивается) поэтому распад и создание. Вся химия углерода на этом построена 1000 орган соединений создает у других хим элементов этого свойства нет. Иммунная система делает накладку(интерференция)с помощью энергии интерферонов пытаясь разрушить цепочку РНК вируса.Надо помочь организму но не вакциной(вирус быстро мутирует)
Читайте также:
