
Тип симметрии вируса гриппа
Основной компонент вириона – капсид(белковая оболочка), кот-й содержит внутри нуклеиновую к-ту. Нуклеокапсид – нуклеиновая к-та, окружённая капсидом.
Капсиды построены из белковых субъединиц (капсомеров). Капсомер – молекула белка. У многих вирусов, кроме нуклеиновой к-ты, есть ещё специальные ферменты.
По типу строения вирионов выделяют:
1) спиральный тип симметрии – вирусы гриппа, парагриппа и др;
2) квазисферический – кубический или икосаэдральный тип симметрии;
3) смешанный у Т-чётных бактериофагов – головка в виде многогранников, хвост – спиралью.
Тип симметрии определяется только нуклеокапсидом, суперкапсид при этом не учитывают. Суперкапсид – это дополнительная оболочка, или пеплос.
20. Изменения каких наружных белков вируса гриппа приводят к появлению пандемических вариантов и почему?
Типовые Аг вирусов гриппа А – гемагглютинин и нейраминидаза, на сочетании этих белков основана классификация вирусов гриппа. Вариации этих белков приводят к появлению новых серовариантов. Антигенные различия среди вирусов типов А, В, С определяют различия в структурах нуклеопротеидных (NP) и матриксных (М) белков. Вирусы типа А имеют групповой (S) Аг, выявляемый в РТГА.
Изменения антигенной структуры происходят 2 путями:
1) антигенный дрейф – точечные мутации, структура Аг меняется незначительно. В большей степени меняется структура гемагглютинина. Дрейф развивается в динамике эпидемического процесса и снижает специфичность иммунных реакций;
2) антигенный шифт – вызывает появление нового антигенного варианта вируса. Происходит в рез-те генетической рекомбинации между штаммами вируса, исчерпавшего свои возможности. Каждые 10-20 лет обновляется популяция людей, но иммунная прослойка исчезает, что приводит к формированию пандемии. Смотри 21.
21. Как устроен вирус гриппа?
Гемагглютинин способствует проникновению вирусов в клетки путём слияния с мембраной клетки. Нейраминидаза распознаёт и взаимодействует с рецепторами.
7 сегментов генома кодируют структурные белки, 8-й – неструктурные, кот-е существуют только в инфицированных клетках. Основные белки:
· матриксный (М-белок) – выполняет защитную ф-ю – окружает геном;
· нуклеопротеидный (NP) – выполняет регуляторную и структурную ф-и.
22. Какая вакцина - против полиомиелита?
В нашей стране для создания активного коллективного иммунитета против полиомиелита используют живую вакцину Сэбина.
23. В каком возрасте делают прививки от полиомиелита?
В России – 3-кратная иммунизация детей с 3-месячного возраста и последующая 4-кратная ревакцинация (до 3-х лет).
24. Как выделить от больного гриппом возбудителя и установить его серотип?
Материал для исследования – смывы и мазки из носоглотки, мазки-отпечатки из носа и кровь.
Экспресс-диагностика – определение Аг вируса в мазках-отпечатках из носа в РИФ и ИФА.
Выделение возбудителя проводят заражением 10-суточного куриного эмбриона. Типовая принадлежность вирусов определяется в РСК, подтип гемагглютинина – в РТГА, подтип нейраминидазы – в реакции ингибирования активности фермента.
25. Как устроен вирус полиомиелита, каковы способы заражения?
Вирион d = 22-30 нм. Вирусы мелкие РНК-геномные. Нуклеокапсид организован по типу кубической симметрии. Суперкапсида нет. Вирусы кислотоустойчивы, стабильны при низких pH, что позволяет им выживать в желудке, отсутствие суперкапсида делает их нечувствительными к д-ю желчных кислот.
Путь заражения фекально-оральный, через загрязнённую воду.
26. Какие методы применяются для диагностики вирусного гепатита А?
Маркеры репликации вируса – АТ (IgM и IgG) к Аг вирусного гепатита А и вирусная РНК. Эти маркеры определяют в ИФА и радиоиммунном анализе, а также в реакции непрямой гемагглютинации. Наличие вирусной РНК можно установить методом ПЦР и методом молекулярной гибридизации.
27. Какая р-я прим-ся для обнаружения HBs-Аг вируса гепатита В у б-х и носителей?
Впервые этот Аг выделен из крови австралийского аборигена, поэтому и называется австралийским. HBs-Аг появляется в крови через 1.5 мес после инфицирования, циркулирует постоянно.
Для выявления HBs-Аг применяют ИФА и РНГА. На наличие свежей инфекции указывают высокие титры HBs-Аг и IgM.
HBs-Аг можно обнаружить в сыворотке крови и слюне больного.
При обнаружении в сыворотке крови ДНК вирусного гепатита В говорят о виремии.
С помощью ПЦР ДНК можно обнаружить за 2-3 нед до появления клиники заболевания.
При вирусоносительстве имеет значение выявление ДНК (методом ПЦР).
28. Какие вакцины используют для создания иммунитета против гепатита В?
Пассивная иммунизация показана лицам, контактировавшим с патологическим материалом и носителями HВs-Аг. Разработаны 2 типа вакцин:
1. Первые вакцины готовят из плазмы пациентов, кот-я содержит Аг вируса гепатита В.
2. Рекомбинантные вакцины – получены методом генной инженерии на культурах пекарских дрожжей.
Массовая иммунизация – взрослые получают 2 дозы в течение месяца и 1 – через 6 мес. Дети получают 1 дозу при рождении и далее через 2 мес и к концу 1-го года жизни.
29. Что такое возбудитель D (дельта)-гепатита?
Это – дефектный РНК-геномный вирус рода Deltavirus. Его выделяют только от пациентов, инфицированных вирусом гепатита В. Моноинфекция вирусом гепатита D невозможна.
Вирионы сферические, d = 35 нм. Геном – 1 нить кольцевой молекулы РНК (близок к вироидам). Суперкапсид вируса гепатита D содержит значительное кол-во HВs-Аг вируса гепатита В. Пути передачи – парентерально и вертикально (от матери к плоду).
30. Чем характеризуется возбудитель гепатитаА?
Гепатит А – болезнь Боткина, известна с древности. Вирус выделен в 1973 г.
Вирионы сферические, d = 25 нм. Геном – несегментированная молекула +РНК. Нуклеокапсид организован по типу кубической симметрии, капсомеры состоят из 4-х белков. Суперкапсида нет. Пути передачи – фекально-оральный.
Вирус устойчив в окружающей среде, при 21° сохраняется несколько недель. Погибает при 85°. Хорошо переносит низкие t, устойчив к хлору (вода).
31. Как устроен вирус гепатита В? Обратная транскриптаза.
Впервые выделен в 1970 г.
Вирион сферический, d = 42 нм, имеет суперкапсид. Геном образован неполной (1 нить короче) 2-нитевой кольцевой молекулой ДНК. С короткой +нитью связан праймерный белок.
Вирус гепатита В не способен размножаться in vitro.
Вирионы содержат полимеразу, способную транскрибировать ДНК в РНК.
Затем -ДНК используется как матрица для транскрипции +ДНК, но этот процесс остаётся незавершённым и опять образуется двойная молекула с неполной +цепью.
Затем образуется оболочка, содержащая поверхностный Аг (HВs-Аг).
32. Какие мех-мы лежат в основе противовирусного иммунитета?
Основные мех-мы защиты направлены на ограничение и подавление вирусной репродукции в клетках (при антибактериальном иммунитете – разрушение возбудителя).
Орг-м человека невосприимчив к возбудителям заболеваний животных.
Неспецифические факторы противовирусного иммунитета:
1) клеточная ареактивность – отсутствие клеток, способных поддерживать репродукцию вирусов;
2) термолабильные вируснейтрализующие β-ингибиторы – липопротеиды сыворотки крови, способные связывать вирус. Комплекс непрочный, распадается через 2 часа под д-ем трипсина, но это сопровождается необратимой инактивацией вируса;
3) t° тела – при ↑ t° - задержка и подавление репродукции вируса;
4) фагоцитирование - вирусы слабо фагоцитируются и не разрушаются ферментами. Номакрофаги фагоцитируют клетки, поражённые вирусом;
5) интерференция вирусов – один вирус подавляет репродукцию другого в клетке-хозяине. Но это не всегда!
6) интерферон – ингибитор репродукции вирусов. Это гликопротеин: α, β, γ, I типа и II типа. Интерферон нарушает трансляцию вирусной РНК рибосомами клетки-хозяина и прекращает синтез белка.
Специфические факторы: АТ – представлены иммуноглобулинами.
33. Как распознают Т-киллеры клетки, инфицированные вирусом?
Т-клетки лизируют клетки-мишени, инфицированные вирусами; распознают чужеродный вирусный Аг в комплексе с молекулой МНС I на мембране клетки-мишени.
Индукция цитотоксических св-в Т-клетки происходит под д-ем 2-х сигналов:
1) взаимодействие между поверхностной молекулой CD8 лимфоцита и комплексом эпитоп-молекула МНС I на клетке-мишени;
2) интерлейкины, секретируемые макрофагами и Т-клетками.
Цитотоксический эффект Т-киллеров регулируется особыми белками – перфоринами. Нарушение осмотического баланса с внеклеточной средой приводит к гибели клетки.
Т-киллеры распознают чужеродные Аг с помощью 2 типов мембранных гликопротеинов: Т-клеточных рецепторов и CD3.
34. Какую роль играют вирусы Коксаки и ЕСНО?
Вирусы Коксаки сходны с вирусами полиомиелита, но перекрёстно не реагируют. Делятся на 2 группы: А –вызывают диффузный миозит с некрозом поперечно-полосатых мышц; В –поражение ЦНС,некроз скелетных мышц, воспаление селезёнки. Пути передачи – фекально-оральный.
Оглавление
1. Особенности строения вирусов. 6
2. Нуклеиновые кислоты вирусов. 11
2.1. Вирусные ДНК.. 14
2.2. Вирусные РНК.. 16
3. Белки, липиды и углеводы вирусов. 22
3.1. Белки вирусов. 22
3.2. Липиды вирусов. 27
3.3. Углеводы вирусов. 28
Список литературы.. 32
Введение
Вирусы — это организмы, не способные существовать и размножаться самостоятельно. Они относятся к царству Vira.
В определении вируса подчеркивается особая природа их паразитизма, который можно назвать паразитизмом на генетическом уровне. Тот факт, что вирусы способны выживать и размножаться только внутри других клеток, объясняется не отсутствием собственной клеточной организации, а их потребностью в поступлении готовых источников питания. Если бактерии обладают способностью расти и размножаться на искусственных питательных средах, то вирусы, напротив, как настоящие клеточные паразиты, полностью зависят от обмена веществ в клетке-хозяине. Сейчас уже доказано, что отношение вирус—хозяин не ограничивается лишь питанием, а носит более сложный характер[1].
Когда стали возможны современные методы исследования, с помощью электронного микроскопа удалось выявить детали структуры вирусов.
Изучение вирусов началось в 1892 году, когда Д.И. Ивановский открыл вирус табачной мозаики. Вскоре, в 1898 году, была доказана вирусная природа ящура, а в 1917 году - были открыты бактериофаги. Вирусы сначала считали микроорганизмами, а бактериофаги - и вовсе разновидностью ферментов. В 60-х годах благодаря развивающейся молекулярной биологии, было введено понятие вириона - единицы вируса, и стало понятно, что вирусы не являются организмами. В 1962 году на 1-м Международном симпозиуме было сформулировано основное отличие вирусов от других живых организмов: генетический материал вирусов - или ДНК, или РНК, а организмы имеют оба типа нуклеиновых кислот. Другое главное отличие - отсутствие у вирусов собственных систем синтеза белка[2].
Представители царства Вира не только не являются организмами, к ним неприменимы основные таксонометрические единицы биологии: особь, популяция, вид. Сначала казалось, что единица вируса - вирион - белковый кокон, с заключенной внутри вирусной ДНК или РНК. Но выяснилось, что один вирион содержит не все гены вируса, а лишь их фрагменты. Синтез новых вирусов начинается в зараженной клетке, когда в нее проникнут вирионы со всеми фрагментами генотипа - их может быть от 2 до 28. Потому нельзя сказать, что вирион - "вирусная особь". Но даже множество вирионов не образуют вирусную популяцию. Рядом с ними всегда будут так называемые дефектные вирусы и вирусы-саттелиты, - вирусы паразитирующие на другом вирусе. Дефектные вирусы размножаются в клетке только при наличии полноценного вируса-помощника (используя его гены, если дефект самого вируса - дефект гена полимеразы, или используя его белки - если у него дефект гена внутренних или оболочечных белков). При смешанных инфекциях генотип одного вируса находится в оболочке другого. Другой тип паразитирующих вирусов - саттелиты. Они используют полноценный неродственный вирус, и при репликации новый саттелит всегда имеет дефект в генотипе, не позволяющий ему самостоятельно размножаться[3].
Есть разновидности вирусов, которые всегда находятся в клетке - плазмиды,оничасто выполняют полезные функции в клетках бактерий (например, синтезируют токсины, убивающие насекомых, или ферменты, разрушающие антибиотики), их репликация обеспечивается клеткой, а их гены не кодируют синтез белков. Прионы - возбудители спогиформных энцефалопатий (болезнь куру, болезнь Крейтцфельда-Якоби) - это результат выхода из-под контроля генов, кодирующих белки, в результате чего поражаются нервные клетки.
Целью данной работы является изучение биохимической структуры вирусов.
Для достижения поставленной цели в работе рассмотрим следующие задачи:
1. изучим общее строение вирусов;
2. на основе общего строения вирусов рассмотрим более детально нуклеиновый состав вирусов; строение белков, липидов и углеводов.
Особенности строения вирусов
Вирусы – это мельчайшие микробы (их размеры колеблются от 12 до 500 нанометров), не имеющие клеточного строения, белок-синтезирующей системы, содержащие только один тип нуклеиновой кислоты (ДНК или РНК).
Важнейшими отличительными особенностями вирусов являются следующие[4]:
1. Они содержат в своем составе только один из типов нуклеиновых кислот: либо рибонуклеиновую кислоту (РНК), либо дезоксирибонуклеиновую (ДНК), - а все клеточные организмы, в том числе и самые примитивные бактерии, содержат и ДНК, и РНК одновременно.
2. Не обладают собственным обменом веществ, имеют очень ограниченное число ферментов. Для размножения используют обмен веществ клетки-хозяина, ее ферменты и энергию.
3. Могут существовать только как внутриклеточные паразиты и не размножаются вне клеток тех организмов, в которых паразитируют.
Вирусы состоят из следующих основных компонентов:
1. Сердцевина - генетический материал (ДНК либо РНК), который несет информацию о нескольких типах белков, необходимых для образования нового вируса.
2. Белковая оболочка, которую называют капсидом (от латинского капса - ящик). Она часто построена из идентичных повторяющихся субъединиц - капсомеров. Капсомеры образуют структуры с высокой степенью симметрии.
3. Дополнительная липопротеидная оболочка. Она образована из плазматической мембраны клетки-хозяина и встречается только у сравнительно больших вирусов (грипп, герпес).
Капсид и дополнительная оболочка несут защитные функции, как бы оберегая нуклеиновую кислоту. Кроме того, они способствуют проникновению вируса в клетку. Полностью сформированный вирус называется вирионом.
Вирусы способны размножаться только в клетках других организмов. Вне клеток организмов они не проявляют никаких признаков жизни. Многие из них во внешней среде имеют форму кристаллов. Размеры вирусов колеблются в пределах от 20 до 300 нм в диаметре[5].
Хорошо изучен вирус табачной мозаики, имеющий палочковидную форму и представляющий собой полый цилиндр. Стенка цилиндра образована молекулами белка, а в его полости расположена спираль РНК (рис. 1). Белковая оболочка защищает нуклеиновую кислоту от неблагоприятных условий внешней среды, а также препятствует проникновению ферментов клеток к РНК и ее расщеплению.

Рис. 1. Схема строения вируса (а) и бактериофага (б); 1— нуклеиновая кислота; 2 — белковая оболочка; 3 — полый стержень; 4 — базальная пластинка; 5 — отростки (нити).
Молекулы вирусной РНК могут самовоспроизводиться. Это означает, что вирусная РНК является источником генетической информации и одновременно иРНК. Поэтому в пораженной клетке в соответствии с программой нуклеиновой кислоты вируса на рибосомах клетки хозяина синтезируются специфические вирусные белки и осуществляется процесс самосборки этих белков с нуклеиновой кислотой в новые вирусные частицы. Клетка при этом истощается и погибает. При поражении некоторыми вирусами клетки не разрушаются, а начинают усиленно делиться, часто образуя у животных, в том числе и человека, злокачественные опухоли.
Поскольку основу всего живого составляют генетические структуры, то и вирусы классифицируют сейчас по характеристике их наследственного вещества - нуклеиновых кислот. Все вирусы подразделяют на две большие группы: ДНК-содержащие вирусы (дезоксивирусы) и РНК-содержащие вирусы (рибовирусы). Затем каждую из этих групп подразделяют на вирусы с двухнитчатой и однонитчатой нуклеиновыми кислотами. Следующий критерий - тип симметрии вирионов (зависит от способа укладки капсомеров), наличие или отсутствие внешних оболочек и т.п.
Ниже в таблице 1 представлена современная классификация вирусов и в качестве примера приведены наиболее известные вирусы.
Классификация вирусов.
Приведенная таблица имеет некоторое сходство с таблицей Менделеева. В ней тоже есть незаполненные места. Так, например, до сих пор неизвестны дезоксивирусы со свойствами 2.2 (однонитчатая ДНК, спиральный тип симметрии) или рибовирусы со свойствами 1.2 (РНК двухнитчатая, смешанный тип симметрии). Может быть, что таких вирусов и нет в природе, а может, их еще не открыли. Совсем недавно рибовирусы со свойствами 1.1.1 не были известны, но затем оказалось, что к ним относятся реовирусы и сходные с ними вирусы раневых опухолей растений. То же самое относится и к дезоксивирусам со свойствами 2.1.1.
Ближайшие годы покажут, реализовала ли природа все возможные схемы строения вирусов, или некоторые из них оказались нежизненными и потому нереализованными.
 |  |
| а | б |
 |  |
| в | г |
Рис. 2. Схематичное изображение расположения капсомеров в капсиде вирусов. Спиральный тип симметрии имеет вирус гриппа - а. Кубический тип симметрии у вирусов: герпеса - б, аденовируса - в, полиомиелита - г.
Из таблицы классификации вирусов видно, что разнообразие в царстве вирусов значительно более выражено, нежели в растительном и животном мире, если за основу взять характеристику генетических структур. В самом деле, все животные и растения - от амебы до человека и от бактерии до цветкового растения - имеют генетический материал в виде двухнитчатой ДНК. У вирусов же генетическим материалом могут быть однонитчатые и двухнитчатые формы обеих нуклеиновых кислот.
Вирусы по праву можно назвать настоящими хозяевами нашей планеты. Где бы ученые не производили исследования, они натыкаются на уже известные вирусы или открывают новые, которые способны выживать при экстремально низких и высоких температурах, в кислотных или щелочных средах. Вирусы обладают уникальной способностью, не свойственной никаким другим живым организмам – хранить генетическую информацию на РНК. Из чего состоят вирусы? Как они внедряются в живую клетку? Как реплицируют свой генетический материал?
Общие сведения о вирусах
Вирус, находящийся вне живой клетки, не проявляет никаких признаков жизни. Такая частица называется вирионом. Вирионы бывают простыми и сложными. Простой состоит из внешней оболочки (капсида) и генетического материала вируса (ДНК или РНК). У сложных вирусов для защиты генома от неблагоприятных факторов есть еще одна оболочка – суперкапсид. В состав капсида входят в основном белки и небольшое количество липидов – холестерина и фосфолипидов. В составе суперкапсида присутствуют также гликопротеины, которые принимают непосредственное участие в проникновении вируса в клетку. Размножаются вирусы путем экспрессии своего генома в зараженной клетке.

Изучим особенности структуры вирусов более подробно.
Строение капсида
Вирионная частица защищена от внешних воздействий плотной оболочкой – капсидом. Это структура, благодаря которой геном вируса защищен от многих повреждений до проникновения в клетку. Помимо белков и липидов, в облочке присутствуют ферменты. Капсид состоит из капсомеров – белковых молекул, которые можно различить в микроскоп.
Геном и белковая оболочка, защищающая его, вместе называются нуклеокапсидом.
Суперкапсид
Простые вирусы защищены от внешних факторов только капсидом, в то время как у сложных вирусов есть дополнительная липопротеиновая оболочка – суперкапсид. Как и капсид, он состоит в основном из белков и липидов.

На поверхности суперкапсидов многих вирусов есть гликопротеидные наросты (пепломеры). Гликопротеины принимают участие в идентификации клеток и связывании вируса с клеткой-мишенью. Затем вирусная оболочка сливается с мембраной хозяина, а тем временем капсид и вирусный геном проникают в клетку и встраиваются в нее. Суперкапсид формируется на этапе сборки вирусных частиц и выхода вируса из клетки.
Функции капсида
Основная функция суперкапсида и капсида – защита генетического материала вируса. А также доставка генома вируса в клетку и взаимодействие с иммунной системой хозяина.
Внешняя оболочка вируса призвана уберечь генетический материал от летальных химических и физических факторов. К ним относятся радиационное излучение, резкие изменения рН или температуры, действие протеолитических и нуклеолитических ферментов.

При поставке генома вируса в клетку капсид (или суперкапсид) связывается с внешними рецепторами клетки-хозяина. На поверхности вирусов, имеющих суперкапсид, за адсорбцию вируса на поверхности клетки хозяина отвечают пепломеры. На поверхности пепломеров некоторых вирусов, например, миксовирусов или вируса гриппа, присутствует белок гемагглютинин, вызывающий агглютинацию эритроцитов. На поверхности суперкапсидных отростков также присутствует нейраминидаза, разрушающая мембрану клетки.
Вирусы без суперкапсидной оболочки как правило проникают через мембрану клетки-хозяина полностью. Остальные вирусы проникают в клетку после слияния внешней оболочки с плазматической мембраной. При этом в цитоплазму клетки попадает только вирусный геном.
Типы симметрии капсида
Поскольку капсид – это сложная структура, состоящая из мелких субъединиц, очевидно, что эти структурные компоненты могут быть организованы по-разному. Именно поэтому существую различные типы симметрии капсидов.
Один из самых распространенных в природе типов капсида – это икосаэдрический. Он характерен для аденовирусов и многих бактериофагов. Капсомеры расположены так, что формируют невыпуклый многогранник с 12 вершинами.
Другой тип капсида – это спиральный. При таком типе симметрии белковые субъединицы уложены как бы по спирали вокруг оси симметрии. Спиральный капсид характерен для вирусов желтухи и табачной мозаики. Такая организация делает вирус палочкообразным.

Смешанный тип симметрии, при котором часть капсида имеет икосаэдрическую форму, а часть спиральную, встречается у бактериофагов.
Геном вируса
В отличие от большинства клеток, имеющих в своем составе и ДНК, и РНК, вирионы содержат только одну нуклеиновую кислоту, которая и образует их геном. Большая часть вирусов человека являются РНК-содержащими. Свойство хранить генетический материал на РНК – уникальная особенность вирусов.
Вирус может содержать одноцепочечную или двуцепочечную нуклеиновую кислоту. Двуцепочечную ДНК в составе генома имеют герпесвирусы и аденовирусы, одноцепочечную парвовирусы, двуцепочечную РНК ротавирусы, одноцепочечную РНК астровирусы.
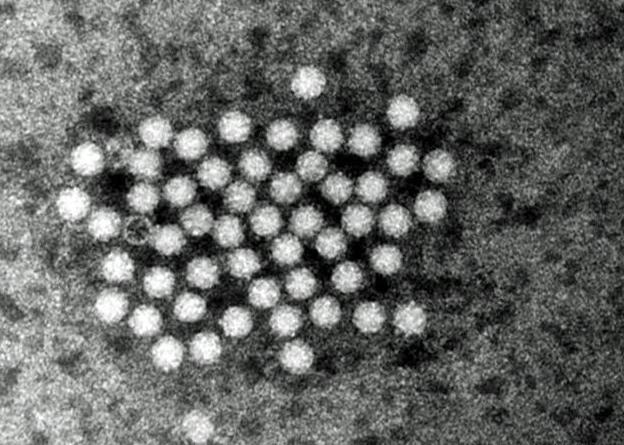
Некоторые вирусы, например, ВИЧ, имеют в составе фермент, который позволяет строить ДНК на матрице РНК – обратную транскриптазу.
Формы вирусов
Вирусы очень разнообразны, как по своему химическому составу, так и по форме. Большинство вирусов (герпес, оспа), имеют сферическую форму. Капсид вируса в этом случае имеет икосаэдрическую симметрию. Распространены также палочковидные вирусы (желтуха, табачная мозаика). Встречаются пулевидные вирусы (вирус бешенства). Необычными являются астровирусы, имеющие звездчатую форму.
Медицинский эксперт статьи


Вирус гриппа А - вирион, который имеет сферическую форму и диаметр 80-120 нм, его молекулярная масса 250 МД. Геном вируса представлен однонитевой фрагментированной (8 фрагментов) негативной РНК с общей м. м. 5 МД. Тип симметрии нуклеокапсида спиральный. Вирус гриппа имеет суперкапсид (мембрану), содержащий два гликопротеида - гемагглютинин и нейраминидазу, которые выступают над мембраной в виде различных шипов. Гемагглютинин имеет структуру тримера с м. м. 225 кД; м. м. каждого мономера 75 кД. Мономер состоит из меньшей субъединицы с м. м. 25 кД (НА2) и большей - с м. м. 50 кД (НА1).
Основные функции гемагглютинина:
- распознает клеточный рецептор - мукопептид, имеющий N-ацетилнейрами-новую (сиаловую) кислоту;
- обеспечивает слияние мембраны вириона с мембраной клетки и мембранами ее лизосом, т. е. отвечает за проникновение вириона в клетку;
- определяет пандемичность вируса (смена гемагглютинина - причина пандемий, его изменчивость - эпидемий гриппа);
- обладает наибольшими протективными свойствами, отвечая за формирование иммунитета.
У вирусов гриппа А человека, млекопитающих и птиц обнаружено 13 различающихся по антигену типов гемагглютинина, которым присвоена сквозная нумерация (отН1доН13).
Нейраминидаза (N) является тетрамером с м. м. 200-250 кД, каждый мономер имеет м. м. 50-60 кД. Ее функции:
- обеспечение диссеминации вирионов путем отщепления нейраминовой кислоты от вновь синтезированных вирионов и мембраны клетки;
- совместно с гемагглютинином определение пандемических и эпидемических свойств вируса.
У вируса гриппа А обнаружено 10 различных вариантов нейраминидазы (N1-N10).
Нуклеокапсид вириона состоит из 8 фрагментов вРНК и капсидных белков, образующих спиралевидный тяж. На З'-концах всех 8 фрагментов вРНК имеются одинаковые последовательности из 12 нуклеотидов. 5'-концы каждого фрагмента также имеют одинаковые последовательности из 13 нуклеотидов. 5'- и 3'-концы частично комплементарны друг другу. Это обстоятельство, очевидно, позволяет осуществлять регуляцию транскрипции и репликации фрагментов. Каждый из фрагментов транскрибируется и реплицируется самостоятельно. С каждым из них прочно связаны четыре капсидных белка: нуклеопротеид (NP), он выполняет структурную и регуляторную роль; белок РВ1 - транскриптаза; РВ2 - эндонуклеаза и РА - репликаза. Белки РВ1 и РВ2 обладают основными (щелочными) свойствами, а РА - кислотными. Белки РВ1, РВ2 и РА образуют полимер. Нуклеокапсид окружен матриксным белком (М1-белком), который играет ведущую роль в морфогенезе вириона и защищает вирионную РНК. Белки М2 (кодирует одна из рамок считывания 7-го фрагмента), NS1 и NS2 (кодируются восьмым фрагментом вРНК, который имеет, как и седьмой фрагмент вРНК, две рамки считывания) синтезируются в ходе репродукции вируса, но в его структуру не входят.

[1], [2], [3], [4], [5]
Жизненный цикл вируса гриппа А
Вирус гриппа абсорбируется на мембране клетки происходит благодаря взаимодействию его гемагглютинина с мукопептидом. Затем вирус проникает в клетку с помощью одного из двух механизмов:
- слияние мембраны вириона с мембраной клетки или
- по пути окаймленная ямка - окаймленный пузырек - эндосома - лизосома - слияние мембраны вириона с мембраной лизосомы - выход нуклеокапсида в цитозоль клетки.
Второй этап "раздевания" вириона (разрушение матриксного белка) происходит на пути к ядру. Особенность жизненного цикла вируса гриппа заключается в том, что для транскрипции его вРНК необходима затравка. Дело в том, что вирус не может сам синтезировать "шапочку", или кэп (англ. cap) - особый участок на 5'-конце мРНК, состоящий из метилированного гуанина и 10- 13 прилежащих нуклеотидов, который необходим для распознавания мРНК рибосомой. Поэтому он с помощью своего белка РВ2 откусывает шапочку от клеточной мРНК, а так как синтез мРНК в клетках происходит только в ядре, вирусная РНК должна обязательно проникнуть сначала в ядро. Она проникает в него в виде рибонуклеопротеида, состоящего из 8 фрагментов РНК, связанных с белками NP, PB1, РВ2 и РА. Теперь жизнь клетки полностью подчиняется интересам вируса, его репродукции.
В ядре на вРНК синтезируются три типа вирус-специфических РНК: 1) позитивные комплементарные РНК (мРНК), используемые в качестве матриц для синтеза вирусных белков; они содержат на 5'-конце шапочку, отщепленную от 5'-конца клеточной мРНК, а на З'-конце - поли-А-последовательность; 2) полноразмерная комплементарная РНК (кРНК), которая служит матрицей Для синтеза вирионных РНК (вРНК); на 5'-конце кРНК шапочки нет, на З'-конце отсутствует поли-А-последовательность; 3) негативная вирионная РНК (вРНК), являющаяся геномом для вновь синтезированных вирионов.
Немедленно, еще до завершения синтеза, вРНК и кРНК вступают в ассоциацию с капсидными белками, которые поступают в ядро из цитозоля. Однако в состав вирионов включаются только рибонуклеопротеиды, связанные с вРНК. Рибонуклеопротеиды, содержащие кРНК, не только не попадают в состав вирионов, но даже не покидают ядро клетки. Вирусные мРНК поступают в цитозоль, где и транслируются. Вновь синтезированные молекулы вРНК после ассоциации с капсидными белками мигрируют из ядра в цитозоль.
[6], [7], [8], [9], [10], [11], [12], [13]
Белки NP, PB1, РВ2, РА и М синтезируются на свободных полирибосомах. Белки NP, PB1, РВ2 и РА после синтеза из цитозоля возвращаются в ядро, где и связываются с вновь синтезированной вРНК, а затем в виде нуклеокапсида возвращаются в цитозоль. Белок матриксный после синтеза движется к внутренней поверхности клеточной мембраны, вытесняя из нее в этом участке клеточные белки. Белки Н и N синтезируются на рибосомах, связанных с мембранами эндоплазматического ретикулума, транспортируются по ним, подвергаясь гликозилированию, и устанавливаются на внешней поверхности клеточной мембраны, образуя шипы как раз напротив белка М, расположенного на ее внутренней поверхности. Белок Н подвергается в ходе процессинга разрезанию на НА1 и НА2.
Заключительный этап морфогенеза вириона контролируется М-белком. С ним взаимодействует нуклеокапсид; он, проходя через мембрану клетки, покрывается вначале М-белком, а затем клеточным липидным слоем и суперкапсидными гликопротеидами Н и N. Жизненный цикл вируса занимает 6-8 ч и завершается отпочковыванием вновь синтезированных вирионов, которые способны атаковать другие клетки ткани.
Устойчивость вируса во внешней среде невелика. Он легко разрушается при нагревании (при 56 °С в течение 5-10 мин), под действием солнечного и УФ-света и легко обезвреживается дезинфицирующими веществами.
[14], [15], [16], [17], [18], [19], [20], [21], [22], [23]
Патогенез и симптомы гриппа А
Инкубационный период при гриппе короткий - 1-2 сут. Вирус размножается в эпителиальных клетках слизистой оболочки дыхательных путей с преимущественной локализацией в области трахеи, что клинически проявляется в виде сухого мучительного кашля с болями по ходу трахеи. Продукты распада пораженных клеток попадают в кровь, вызывают сильную интоксикацию и повышение температуры тела до 38-39 °С. Повышение проницаемости сосудов, обусловленное повреждением клеток эндотелия, может стать причиной патологических изменений в различных органах: точечных кровоизлияний в трахее, бронхах, а иногда и отека мозга с летальным исходом. Вирус гриппа оказывает угнетающее действие на кроветворение и иммунную систему. Все это может приводить к вторичным вирусным и бактериальным инфекциям, которые осложняют течение болезни.
Постинфекционный иммунитет
Прежние представления о том, что после перенесенного гриппа остается слабый и кратковременный иммунитет, опровергнуты после возвращения вируса H1N1 в 1977 г. Этот вирус вызывал заболевание главным образом у людей не старше 20 лет, т. е. у тех, кто не болел им раньше, до 1957 г. Следовательно, постинфекционный иммунитет достаточно напряженный и продолжительный, но имеет выраженный типоспецифический характер.
Главная роль в формировании приобретенного иммунитета принадлежит вирус-нейтрализующим антителам, блокирующим гемагглютинин и нейраминидазу, а также секреторным иммуноглобулинам IgAs.
Эпидемиология гриппа А
Источник инфекции - человек, больной или носитель, редко животные (домашние и дикие птицы, свиньи). Заражение от людей происходит воздушно-капельным путем, инкубационный период очень короткий (1-2 сут.), поэтому эпидемия распространяется очень быстро и может при отсутствии коллективного иммунитета перерасти в пандемию. Иммунитет - основной регулятор эпидемий гриппа. По мере нарастания коллективного иммунитета эпидемия идет на убыль. Вместе с тем вследствие формирования иммунитета происходит отбор штаммов вируса с измененной антигенной структурой, прежде всего гемагглютинина и нейраминидазы; эти вирусы продолжают вызывать вспышки до тех пор, пока и к ним не появятся антитела. Такой антигенный дрейф и поддерживает непрерываемость эпидемии. Однако у вируса гриппа А обнаружена еще одна форма изменчивости, получившая название шифта, или сдвига. Она связана с полной сменой одного типа гемагглютинина (реже - и нейраминидазы) на другой.
Все пандемии гриппа были вызваны вирусами гриппа А, претерпевшими шифт. Пандемия 1918 г. была вызвана вирусом с фенотипом H1N1 (погибло около 20 млн человек), пандемия 1957 г. - вирусом h3N2 (переболело более половины населения мира), 1968 г. - вирусом H3N2.
Для объяснения причин резкой смены типов вирусов гриппа А предложены две основные гипотезы. Согласно гипотезе А. А. Смородинцева, вирус, исчерпавший свои эпидемические возможности, не исчезает, а продолжает циркулировать в коллективе без заметных вспышек или длительно персистировать в организме человека. Через 10-20 лет, когда появится новое поколение людей, не имеющих иммунитета к этому вирусу, он становится причиной новых эпидемий. В пользу этой гипотезы говорит тот факт, что вирус гриппа А с фенотипом H1N1, исчезнувший в 1957 г., когда его вытеснил вирус h3N2, вновь появился после 20-летнего отсутствия в 1977 г.
По другой гипотезе, развиваемой и поддерживаемой многими авторами, новые типы вируса гриппа А возникают вследствие реассоциации геномов между вирусами гриппа человека и птиц, между вирусами гриппа птиц, между вирусами гриппа птиц и млекопитающих (свиньи), чему способствует сегментарная структура вирусного генома (8 фрагментов).
Таким образом, у вируса гриппа А есть два пути изменения генома.
Точечные мутации, обусловливающие антигенный дрейф. Им подвержены, прежде всего, гены гемагглютинина и нейраминидазы, особенно у вируса H3N2. Благодаря этому вирус H3N2 за период с 1982 по 1998 г. вызвал 8 эпидемий и сохраняет эпидемическое значение до сих пор.
Реассоциация генов между вирусами гриппа человека и вирусами гриппа птиц и свиней. Считается, что именно реассоциация геномов вирусов гриппа А с геномами вируса гриппа птиц и свиней - главная причина возникновения пандемических вариантов этого вируса. Антигенный дрейф позволяет вирусу преодолевать существующий у людей иммунитет. Антигенный шифт создает новую эпидемическую ситуацию: к новому вирусу у большинства людей иммунитета нет, и возникает пандемия гриппа. Возможность такой реассоциации геномов вирусов гриппа А доказана экспериментально.
Установлено, что эпидемии гриппа у людей вызывают вирусы типа А только 3 или 4 фенотипов: H1N1 (H0N1); h3N2; H3N2.
Однако существенную угрозу для человека предсталяет и куриный (птичий) вирус. Вспышки куриного гриппа наблюдались неоднократно, в частности куриный вирус H5N1 вызвал миллионную эпизоотию среди домашних и диких птиц с 80- 90 %-ной летальностью. От кур заражались и люди; так в 1997 г. от кур заразилось 18 человек, треть из них погибла. Особенно крупная вспышка наблюдалась в январе-марте 2004 г. Она охватила почти все страны Юго-Восточной Азии и один из штатов США и нанесла огромный экономический ущерб. От кур заразилось и погибло 22 человека. Для ликвидации этой вспышки были предприняты самые жесткие и решительные меры: строгий карантин, ликвидация всего поголовья птиц во всех очагах, госпитализация и изоляция больных и всех людей с повышенной температурой, а также лиц, находившихся в контакте с больными, запрет импорта куриного мяса из указанных выше стран, строгий медицинский и ветеринарный надзор за всеми пассажирами и транспортными средствами, прибывающими из этих стран. Широкого распространения гриппа среди людей не произошло потому, что не было реассоциации генома вируса куриного гриппа с геномом вируса гриппа человека. Однако опасность такой реассоциации остается реальной. Это может привести к появлению нового опасного пандемического вируса гриппа человека.
В названии выявляемых штаммов вирусов гриппа указывают серотип вируса (А, В, С), вид хозяина (если им не является человек), место выделения, номер штамма, год его выделения (последние 2 цифры) и фенотип (в круглых скобках). Например: "А/Сингапур/1/57 (h3N2), А/утка/СССР/695/76 (H3N2)".
[24], [25], [26], [27], [28], [29], [30], [31], [32], [33], [34]
Читайте также:
