
Ssrna вирусы животных общие характеристики





Описание книги "Общая вирусология с основами таксономии вирусов позвоночных"
Описание и краткое содержание "Общая вирусология с основами таксономии вирусов позвоночных" читать бесплатно онлайн.
Общая вирусология с основами таксономии вирусов позвоночных
Вирусология – наука о морфологии, физиологии, генетике, экологии и эволюции вирусов. Медицинская и ветеринарная вирусология исследует вирусы-паразиты человека и животных, их роль в этиологии и патогенезе инфекционных и опухолевых болезней, разрабатывает специальные методы диагностики, способы этиотропной терапии и специфической профилактики.
Вирусы являются одной из наиболее распространенных групп живых организмов, которые способны заражать не только практически всех представителей флоры и фауны, но и микроорганизмы. Эффективность борьбы со многими вирусами не имеет положительных результатов, так как вирусы не только имеют природный резервуар в окружающей среде, но и постоянно изменяются (мутируют), в связи с чем снижается эффективность проведения вакцинопрофилактики. Одним из наиболее ярких примеров является неэффективная многолетняя борьба с вирусом иммунодефицита человека.
Своевременность и точность постановки диагноза вирусного инфекционного заболевания позволяет не только осуществить эффективное лечение, направленное на уничтожение вирусного агента, но и провести профилактику дальнейшего распространения данного заболевания с учетом индивидуальных особенностей вириона. Например, в XX веке повсеместно было ликвидировано такое заболевание как оспа по средствам проведения массовой вакцинации населения.
В данном учебном пособии подробно рассмотрены вопросы основ классификации (таксономии) вирусов позвоночных, международный код классификации и номенклатуры вирусов, характеристики семейств и родов ДНК и РНК-содержащих вирусов, диагностики вирусных заболеваний и методы выделения и очистки вирусных препаратов.
Обозначения и сокращения
AGE – агарозный гель
ANU – Australian National University
ATCC – American Type Culture Collection
bр – (base pair) пара оснований
ds – (double-stranded) двуспиральный
dsDNA – двуспиральная ДНК
dsDNA-RT – двуспиральная ДНК, в репликативном цикле которой имеется этап обратной транскрипции с РНК
dsRNA – двуспиральная РНК
dsRNA – двуспиральная РНК
G (GP) – поверхностный гликопротеин
IN – интеграза
IRES – (internal ribosomal entry site) внутренний сайт входа рибосомы
kb – (kilo base) килобаза (тысяча оснований)
kbp – (kilo base pare) тысяча пар оснований
kDa – (kilo Dalton) килодальтон
MA – (matrix) матрикс
Mr – (relative molar mass) относительная молярная масса
N – (nucleoprotein) нуклеопротеин
NC – (nucleocapsid) нуклеокапсид
NC – (nucleocapsid) нуклеокапсид)
NNS – негативный несегментированный геном РНК
NP – нуклеопротеин
NS – неструктурные белки – это предшественники структурных белков, регуляторные белки и ферменты, обслуживающие процесс внутриклеточной репродукции вируса и не входящие в состав вирусной частицы
NSF – National Science Foundation
NssRNA – односпиральная РНК негативной полярности
nt – (nucleotide) нуклеотид
ORF – (open reading frame) открытая рамка считывания
PAGE – полиакриламидный гель
PR – протеаза
РrР – приемный протеин
RT – (reverse transcriptase) обратная транскриптаза
RT – обратная транскриптаза
sgRNA – (subgenomic RNA) субгеномная РНК
SH – интегративный мембранный протеин
SP – сериновая протеаза
ss – (single-stranded) одноцепочечный
ssDNA – односпиральная ДНК
ssRNA – односпиральная РНК
ssRNA-RT – односпиральная РНК в репликативном цикле которой имеется этап обратной транскрипции
SU – (surface) поверхностный
T – (triangular number) триангулярное число
VPg – (genome-linked protein) геном-ассоциированный протеин
VP – структурные белки, входящие в состав вирусной частицы
БСА – альбумин нормальной сыворотки быка
ДНК – дезоксирибонуклеиновая кислота
ИБ – метод иммуноблотинга
ИД50 – 50 % инфекционная доза для животных или для куриных эмбрионов
иРНК – информационная рибонуклеиновая кислота
ИФА – иммунофлюоресцентный анализ
ИЭМ – иммунная электронная микроскопия
ИЭТ – изоэлектрическая точка
кДНК – клеточная дезоксирибонуклеиновая кислота
КПЧ – клетки почек человека
ЛД50 – 50 % смертельная доза
М – матриксный протеин
ММГ – методы молекулярной гибридизации
мРНК – матричная рибонуклеиновая кислота
МФА – метод иммунофлюоресценции
ПЦР – полимеразная цепная реакция
ПЭГ – полиэтиленгликоль
РБО – реакция бляшкообразования
РВИЭФ – реакция встречного иммуноэлектрофореза
РГА – реакция гемагглютинации
РГАд – реакции гемадсорбции
РЗБО – реакция задержки (нейтрализации) бляшкообразования
РЗЦПД – реакция задержки цитопатического действия вирусов
РИФ – реакция иммунофлюоресценции
РН – реакция нейтрализации
РНК – рибонуклеиновая кислота
РРГ – реакция радиального гемолиза
рРНК – рибосомальная рибонуклеиновая кислота
РСК – реакция связывания комплемента
РСР – папаино-подобная цистеиновая протеаза
РТГА – реакция торможения гемагглютинации
РТГАд – реакция торможения гемадсорбции
СА – (capsid protein) капсидный протеин
СР – (capsid protein) капсидный протеин
СРЕ – (cytopathic effect) ЦПЭ, цитопатический эффект
ТКИД50 – 50 % инфекционная доза для тканевой культуры
ТМ – (transmembrane) трансмембранный
тРНК – транспортная рибонуклеиновая кислота
УФ – ультрафиолет
ФИТЦ – флюоресцеина изотиоцианат
ФЭЧ – фибробласты эмбриона человека
ЦПД – цитопатическому действию
ЭМВ – электронно-микроскопическое выявление
1 История вирусологии, природа и происхождение вирусов
1.1 Открытие вирусов
Вирусология – молодая наука, ее история насчитывает немногим более 100 лет. Начав свой путь как наука о вирусах, вызывающих болезни человека, животных и растений, в настоящее время вирусология развивается в направлениях изучения основных законов современной биологии на молекулярном уровне, основываясь на том, что вирусы являются частью биосферы и важным фактором эволюции органического мира.
История вирусологии необычна тем, что один из ее предметов – вирусные болезни – стал изучаться задолго до того, как были открыты собственно вирусы. Начало истории вирусологии – это борьба с инфекционными заболеваниями и только впоследствии – постепенное раскрытие источников этих болезней. Подтверждением тому служат работы Эдуарда Дженнера (1749-1823 гг.) по предупреждению оспы и работы Луи Пастера (1822-1895 гг.) с возбудителем бешенства.
С незапамятных времен оспа была бичом человечества, унося тысячи жизней. Описания оспенной заразы встречаются в рукописях древнейших китайских и индийских текстов. Первые упоминания об эпидемиях оспы на европейском континенте датируются VI столетием нашей эры (эпидемия среди солдат эфиопской армии, осаждавшей Мекку), после чего наблюдался необъяснимый период времени, когда упоминания об эпидемиях оспы отсутствовали. Оспа снова начала гулять по континентам в XVII веке. Например, в Северной Америке (1617-1619 гг.) в штате Массачусетс погибло 9/10 населения, в Исландии (1707 г.) после эпидемии оспы от 57 тыс. человек осталось только 17 тыс., в г. Истхем (1763 г.) от 1331 жителя осталось 4 человека. В связи с этим, проблема борьбы с оспой стояла очень остро.
Методика предупреждения оспы через прививку, называемая вариоляцией, была известна с давних времен. Упоминания о применении вариоляции в Европе датируются серединой 17-го века со ссылками на более ранний опыт применения в Китае, на Дальнем Востоке, в Турции. Суть вариоляции заключалась в том, что содержимое пустул от пациентов, болевших легкой формой оспы, вносили в маленькую ранку на коже человека, что вызывало легкое заболевание и предупреждало острую форму. Однако при этом сохранялась большая опасность заболевания тяжелой формой оспы и смертность среди привитых достигала 10 %. Дженнер совершил переворот в методике предупреждения оспы. Он первый обратил внимание на то, что люди, переболевшие коровьей оспой, которая протекала легко, впоследствии никогда не болели оспой. 14 мая 1796 г. Дженнер внес в ранку Джеймса Фипса, никогда не болевшего оспой, жидкость из пустул больной коровьей оспой доярки Сары Селмес. На месте искусственной инфекции у мальчика появились типичные пустулы, которые через 14 дней исчезли. Тогда Дженнер внес в ранку мальчика высокоинфекционный материал из пустул больного оспой. Мальчик не заболел. Так зародилась и подтвердилась идея вакцинации (от латинского слова vacca – корова). Во времена Дженнера вакцинация понималась как внесение инфекционного материала коровьей оспы в организм человека с целью предотвращения заболевания натуральной оспой. Термин вакцина применяли к веществу, предохранявшему от оспы. С 1840 г. противооспенную вакцину стали получать заражением телят. Вирус оспы человека был открыт только в 1904 г. Таким образом, оспа – это первая инфекция, против которой была применена вакцина, т. е. первая управляемая инфекция. Успехи в вакцинопрофилактике черной оспы привели к ее искоренению в мировом масштабе.
ВНИМАНИЕ! САЙТ ЛЕКЦИИ.ОРГ проводит недельный опрос. ПРИМИТЕ УЧАСТИЕ. ВСЕГО 1 МИНУТА.
Начало истории вирусологии связано с именем Д.И. Ивановского, который в 1892 г. опубликовал работу по изучению мозаичной болезни табака. Он отметил, что возбудитель – мельчайшее существо, проходит через бактериальные фильтры, не растет на питательных средах, невидим в световом микроскопе.
В 1898 г. Леффлер и Фрош открыли вирус ящура.
В 1901 г. Рид и Кэррол выделили вирус из трупов людей, умерших от желтой лихорадки.
ДўЭррель в 1910 г. обнаружил вирусы бактерий – бактериофаги.
Вирусы широко распространены в природе, окружающей среде и практически вездесущи. Они находятся в воздухе, воде, пище, космосе и в живых организмах, а вирусы бактерий – бактериофаги – в бактериях.
Медицинская вирусология изучает лишь вирусы, патогенные для человека или значимые для медицины (бактериофаги).
Основной задачей медицинской вирусологии является изучение морфологии, физиологии, генетики, экологии и эволюции вирусов и разработка методов диагностики, лечения и профилактики инфекций у человека.
Вирусы – облигатные внутриклеточные паразиты, имеющие собственный геном, структурные белки и ферменты, способные репродуцироваться только в чувствительных к ним клетках человека, животных, растений, бактериях. Это своеобразная форма жизни, биологически активные структуры, которые подчиняются законам эволюции.
Основные свойства вирусов:
- не имеют типичного клеточного строения,
- состоят из белков и одной нуклеиновой кислоты (ДНК или РНК), где закодирована вся генетическая информация вируса,
- не обладают собственными метаболическими и энергетическими системами,
- размножаются в цитоплазме или в ядре клеток с использованием белоксинтезирующих и энергетических систем клетки хозяина, так как являются облигатными внутриклеточными паразитами,
- используют рибосомы клетки хозяина для синтеза собственных белков,
- имеют особый способ размножения – дизъюнктивную (разобщенную) репродукцию: в клетке отдельно синтезируются нуклеиновые кислоты и белки вирусов, а затем происходит сборка их в вирусные частицы.
- могут интегрировать свой геном в геном клетки с образованием провируса,
- вирусы имеют малые размеры (от 15 до 250 нм и более).
- как и другие формы жизни вирусы обладают наследственностью и изменчивостью, сохраняют жизнеспособность при замораживании, высушивании, резистентны к антибиотикам, но чувствительны к высокой температуре.
Кроме типичных вирусов известны необычные инфекционные частицы – прионы и вироиды.
Прионы – белковые инфекционные частицы, которые имеют вид фибрилл размером 10-20х100-200 нм, массу 30 кД, не содержат нуклеиновой кислоты, устойчивы к нагреванию, к действию протеаз, ультрафиолетовых лучей, ультразвука и ионизирующей радиации. Прионы возникают как продукты мутации собственного гена или попадают в организм при употреблении мяса животных, содержащего прионы. Прионы накапливаются в пораженном органе, не вызывая цитопатогенного действия (ЦПД), иммунного ответа и воспалительных реакций. Они могут блокировать или активировать гены человека или животного.
Вироиды – это небольшие молекулы кольцевой суперспирализованной РНК, не содержащие белка, вызывающие заболевания у растений, возможно и у млекопитающих.
Классификация вирусов
В силу своих особенностей вирусы выделены в отдельное надцарство Vira, в котором по типу нуклеиновой кислоты различают рибовирусы и дезоксирибовирусы (табл. 1).
Подцарства делятся на семейства, которые подразделяются на подсемейства и роды. Вид – совокупность вирусов, имеющих почти идентичные геном (ДНК или РНК), свойства и способность вызывать определенный патологический процесс. Названия семейства имеют окончание viridae, подсемейство – virinae, рода – virus.
Признаки, используемые для классификации вирусов: 1) тип нуклеиновой кислоты – ДНК или РНК; 2) их структура (однонитевая, двунитевая, линейная, кольцевая, фрагментированная, нефрагментированная с повторяющимися и инвертированными последовательностями); 3) структура, размеры, тип симметрии, число капсомеров; 4) наличие или отсутствие внешней оболочки (суперкапсида); 5) антигенная структура; 6) феномены генетических взаимодействий; 7) круг восприимчивых хозяев; 8) географическое распространение; 9) внутриядерная или цитоплазматическая локализация; 10) чувствительность к эфиру и детергентам; 11) путь передачи инфекции.
Для определения принадлежности к семейству ретровирусов обязательно учитывается наличие фермента обратной транскриптазы.
Вирусы, вызывающие инфекционные процессы у человека, входят в состав как ДНК-содержащих, так и РНК-содержащих вирусных семейств (см. табл. 1).
Таблица 1.
Классификация и некоторые свойства вирусов
| Семейство вирусов | Тип нуклеиновой кислоты | Размер вириона, нм | Наличие суперкапсида | Типовые представители |
| РНК-геномные вирусы | ||||
| Arenaviridae Аренавирусы | фрагментированная, однонитчатая | 50-300 | + | Вирусы Ласса, Мачупо |
| Bunyaviridae Буньявирусы | фрагментированная, однонитчатая, кольцевая | 90-100 | + | Вирусы геморрагических лихорадок и энцефалитов |
| Caliciviridae Калицивирусы | однонитчатая | 20-30 | - | Вирус гепатита Е, калицивирусы человека |
| Coronaviridae Коронавирусы | однонитчатая (+)РНК | 80-130 | + | Коронавирусы человека |
| Orthomyxoviridae Ортомиксо- вирусы | однонитчатая, фрагментированная (-)РНК | 80-120 | + | Вирусы гриппа |
| Paramyxoviridae Парамиксо- вирусы | однонитчатая, линейная (-)РНК | 150-300 | + | Вирусы парагриппа, кори, эпидемического паротита, РС-вирус |
| Picornaviridae Пикорнавирусы | однонитчатая (+)РНК | 20-30 | - | Вирусы полиомиелита, Коксаки, ЕСНО, гепатита А, риновирусы |
| Reoviridae Реовирусы | двунитчатая РНК | 60-80 | - | Реовирусы |
| Retroviridae Ретровирусы | однонитчатая РНК | 80-100 | + | Вирусы рака, лейкоза, саркомы, ВИЧ |
| Togaviridae Тогавирусы | однонитчатая (+)РНК | 30-90 | + | Вирусы лошадиных энцефалитов, краснухи и др. |
| Flaviviridae Флавивирусы | однонитчатая (+)РНК | 30-90 | + | Вирусы клещевого энцефалита, желтой лихорадки, Денге, японского энцефалита, гепатитов С, G |
| Rhabdoviridae Рабдовирусы | однонитчатая (-)РНК | 30-90 | + | Вирус бешенства, вирус везикулярного стоматита |
| Filoviridae Филовирусы | однонитчатая (+)РНК | 200-4000 | + | Вирусы лихорадки Эбола, Марбург |
| ДНК-геномные вирусы | ||||
| Adenoviridae Аденовирусы | линейная, двунитчатая | 70-90 | - | Аденовирусы млекопитающихся и птиц |
| Hepadnaviridae Гепаднавирусы | двунитчатая, кольцевая с однонитчатым участком | 45-50 | + | Вирус гепатита В |
| Herpesviridae Герпесвирусы | линейная, двунитчатая | + | Вирусы простого герпеса, цитомегалии, ветряной оспы, инфекционного мононуклеоза | |
| Papovaviridae Паповавирусы | двунитчатая, кольцевая | 45-55 | - | Вирусы папилломы, полиомы |
| Poxviridae Поксвирусы | двунитчатая с замкнутыми концами | 130-250 | + | Вирус осповакцины, вирус натуральной оспы |
| Parvoviridae Парвовирусы | линейная, однонитчатая | 18-26 | - | Аденоассоциированный вирус |
4.2. Строение вирусов
По строению различают два типа вирусных частиц – простые и сложные. В составе простых вирионов есть ДНК или РНК и белки. У сложных в суперкапсиде содержатся липиды, полисахариды.
Внутренняя структура простых и сложных вируосв сходна, сердцевина вируса – вирусный геном, который содержит от 3 до 100 и более генов.
Морфология и структура вирусов. Простые вирусы имеют одну белковую оболочку – капсид, который состоит из капсомеров – белковых молекул, форма укладки которых определяет тип симметрии. Капсид представлен a-спиральными белками, способными к полимеризации.
Сложные вирусы имеют внешнюю оболочку – суперкапсид, расположенную поверх капсида. В состав суперкапсида входит внутренний белковый слой – М-белок, затем более объемный слой липидов и углеводов, извлеченных из клеточных мембран клетки хозяина. Вирусспецифические гликопротеиды проникают внутрь суперкапсида, образуя фигурные выпячивания (шипы, фибры), которые выполняют рецепторную функцию.
Различают 3 типа симметрии: 1) спиральный, когда капсомеры укладываются по спирали – винтообразная структура нуклеокапсида; 2) кубический (икосаэдрический), когда капсомеры укладываются по граням многогранника (12-20-гранника) – в основе лежит фигура икосаэдра (20-гранника). В зависимости от типа перегруппировки и числа субъединиц число капсомеров будет равным 30, 20, или 12. Вирионы со сложным капсидом, построенным более чем из 60 капсомеров, содержат группы из 5 субъединиц – пентамеры, или из 6 субъединиц – гексамеры; 3) смешанный тип симметрии (у бактериофагов).
Комплекс капсида и генома вируса называют нуклеокапсид. Сложные вирусы имеют суперкапсид (пеплос). Эта поверхностная оболочка вируса, состоит из липидов и белков клеточного происхождения.
Вирусные белки бывают: 1) структурные; 2) неструктурные.
Среди структурных различают: капсидные – входят в состав капсомеров и образуют футляр, защищающий нуклеиновую кислоту; суперкапсидные – это гликопротеиды, которые формируют шипы на поверхности суперкапсида и выполняют: адресную функцию – узнают чувствительную клетку и адсорбируются на ней; прикрепительныебелки, которые взаимодействуют со специфическими рецепторами клетки; белкислияния – способствуют слиянию вирусной и клеточной мембран и приводят к образованию симпластов; геномные – обладают антигенными свойсвами, участвуют во взаимодействии с клеткой.
Среди неструктурных белков различают: предшественники вирусных белков (нестабильные); РНК- и ДНК-полимеразы – участвуют в репликации вирусного генома; регуляторные белки – участвуют в репродукции вируса.
Функции белков:обладают антигенными и иммуногенными свойствами; участвуют в распознавании клетки и взаимодействии с ней; защищают геном от нуклеаз; обеспечивают тип симметрии.
Липиды входят в состав суперкапсида и представляют смесь нейтральных фосфо- и гликолипидов, многие из них – продукты мембраны клеток хозяина.
Они обусловливают инфекционность, чувствительность или устойчивость к эфиру; стабилизируют вирусную частицу.
Углеводы входят в состав гликопротеидов суперкапсида. Углеводы и липиды – составная часть гемагглютинина, который вызывает склеивание эритроцитов и обладает антигенной специфичностью.
Различают вирионные и вирусиндуцированные ферменты вирусов. К вирионным относят ферменты транскрипции и репликации (ДНК и РНК-полимеразы); обратную транскриптазу (у ретровирусов), АТФ-азы, эндо- и экзонуклеазы, нейраминидазы.
К вирусиндуцированным относятся ферменты, о которых имеется только информация в вирусном геноме, а появляются они в клетке. Это РНК-полимеразы тога-, орто-, пикорна- и парамиксовирусов; и ДНК-полимеразы у покс- и герпесвирусов.
Нуклеиновые кислотыобеспечивают наследственные признаки; являются хранителями генетической информации; необходимы для репродукции вирусов, многие из них могут вызывать инфекционный процесс самостоятельно, достаточно их проникновения в клетку.
Вирусная ДНК.Молекулярная масса равна 1,10 6 -1,10 8 дальтон. ДНК может быть одно- или двунитчатой, фрагментированной и сверхспирализованной, линейной или кольцевой, содержит несколько сотен генов. В каждой нити ДНК есть нуклеотидные последовательности, а на концах есть прямые или инвертированные (повернутые на 180 о ) повторы, которые являются маркерами для отличия вирусной ДНК от клеточной. Эти повторы обеспечивают способность ДНК замыкаться в кольцо для последующих репликации, транскрибирования и встраивания в клеточный геном. Генетическая информация инфекционной ДНК транслируется на мРНК в клетке с помощью полимераз.
Вирусная РНКможет быть одно- и двунитчатой, линейной, кольцевой, фрагментированной. У РНК-содержащих вирусов генетическая информация закодирована в РНК таким же кодом, как в ДНК всех других вирусов и клеточных организмов. Вирусные РНК по своему химическому составу не отличаются от РНК клеточного происхождения, но характеризуются разной структурой.
Наряду с типичной для всех РНК однонитевой формой у ряда вирусов имеется двунитевая РНК. В составе однонитевых РНК имеются спиральные участки типа двойной спирали ДНК, образующиеся вследствие спаривания комлементарных азотистых оснований. Вирусы с однонитчатой РНК делятся на 2 группы: (+)РНК (положительный геном) и (-)РНК (отрицательный геном). Вирусная (+)РНК инфекционная и обладает функциями информационной РНК. Она может передовать генетическую информацию на рибосомы, как иРНК. Вирусы с отрицательным геномом не обладают инфекциозностью, т.к. нить (-)РНК выполняет только наследственную функцию и не обладает функцией иРНК. В зараженной клетке на матрице вирусной геномной РНК с помощью фермента транскриптазы осуществляется синтез РНК-комплементарной геному.
Патогенность вирусов обусловлена совокупностью их свойств: способностью проникать в макроорганизм, связываться с клеточными мембранами и проникать в клетку, управлять метаболизмом и белоксинтезирующей функцией клетки, обеспечивать транскрипцию и репликацию собственного генома и осуществлять весь цикл репродукции вирусов. Все эти свойства зависят от генома вирусов и наличия соответствующих структурных белков и ферментов. Репродукция вирусов приводит к развитию патологии: цитопатогенному (разрушающему) действию, развитию воспаления, повреждению различных клеток и тканей.
2.1 Основные семейства
2.3 Методы выделения, культивирования и идентификации вирусов
2.1 Основные семейства
Вирусы отнесены к царству Vira. По типу нуклеиновой кислоты выделяют рибовирусы(РНК-вирусы) и дезоксирибовирусы(ДНК-вирусы). Для вирусов предложены следующие таксономические категории (по восходящей): Вид (Species) → Род (Genus) → Подсемейство (Subfamilia) → Семейство (Familia). Категории подсемейств и родов разработаны не для всех вирусов.
Из более чем 55 семейств вирусов, признанных Международным комитетом по таксономии вирусов, следующие 19 включают вирусы человека и животных: поксвирусы, иридовирусы, вирусы герпеса, аденовирусы, паповавирусы, вирусы гепатита В, парвовирусы, реовирусы, тогавирусы, коронавирусы, парамиксовирусы, рабдовирусы, филовирусы, ортомиксовирусы, буньявирусы, аренавирусы, ретровирусы, пикорнавирусы, калицивирусы.
К числу семейств вирусов исключительно позвоночных относятся Herpesviridae, Adenoviridae, Papovaviridae, Paramyxoviridae, Orthomyxoviridae, Arenaviridae, Coronaviridae.
Размножаться в двух типах хозяев – позвоночных и беспозвоночных (клещи, комары, москиты) способны вирусы семейства Bunyaviridae, роды Alphavirus и Flavivirus семейства Togaviridae, вирусы родов Vesiculovirus и Lyssavirus семейства Rhabdoviridae, род Orbivirus семейства Reoviridae, вирус африканской лихорадки свиней семейства Iridoviridae. Такие вирусы составляют экологическую группу арбовирусов, т. е. вирусов позвоночных, передающихся членистоногими.
Рисунок 18 – Поксвирусы:
А – кирпичеобразная форма вириона; Б – вирус контагиозного моллюска
Вирион включает сердцевину, окруженную гладкой мембраной и слоем равномерно расположенных цилиндрических структур. Снаружи располагаются овальные структуры (белковые тела), окруженные оболочкой с характерной бороздчатой структурой. Геном – двухнитевая молекула ДНК.
Семейство включает вирусы осповакцины и натуральной оспы (род Orthopoxvirus), вирусы узелков доярок – псевдооспы крупного рогатого скота (род Parapoxvirus), вирус контагиозного моллюска (род Molluscipoxvirus) (рисунок 18 Б), вирусы оспы Тана и Яба – оспы обезьян (род Yatapoxvirus).
Вирусы герпеса. Герпесвирусы Herpesviridae [от греч. herpes, ползучее поражение кожи] – группа сравнительно крупных вирусов диаметром 150–200 нм (рисунок 19 А, Б) и кубическим типом симметрии. Геном представлен двухнитевой молекулой ДНК, содержащей короткий (18 %) и длинный (82 %) компоненты.
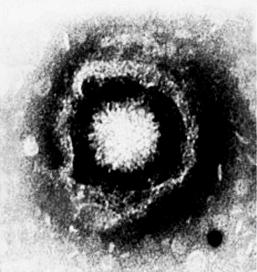
Рисунок 19 – Герпесвирусы:
Суперкапсиды герпесвирусов образованы фрагментами ядерных мембран (созревание дочерних популяций происходит на внутренней мембране ядер зараженной клетки – рисунок 20) и пронизан гликопротеиновыми шипами. Между нуклеокапсидом и суперкапсидом расположен покровный слой – тегумент.
Аденовирусы. Патогенные для человека вирусы включены в состав рода Mastadenovirus (аденовирусы млекопитающих) семейства Adenoviridae. Аденовирусы организованы по принципу кубической симметрии и не имеют суперкапсида. Геном представлен линейной молекулой двухнитевой ДНК. Средний диаметр вириона равен 60–90 нм. Капсид состоит из 252 капсомеров, 240 из них (гексоны) образуют его грани, 12 (пентоны) – полигональные основания и прикреплённые к нему нити (рисунок 21).
Рисунок 21 – Вид аденовируса при электронной микроскопии (А) (˟ 600000 раз) и его модель (Б)
Паповавирусы. Название вирусов указывает на способность вызывать опухолевые трансформации клеток [па(пиллома) + по(лиома) + ва(куолизирующие) вирусы]. Семейство Papovaviridae включает роды Papillomavirus и Polyomavirus (рисунок 22 А, Б), последние приводят к развитию папиллом и полиом у своих хозяев – различных млекопитающих (в том числе и человека). Геном образует кольцевая ДНК; капсид организован по типу кубической симметрии. Средний размер вирионов 45–50 нм.
Рисунок 22 – Папилломавирус (А) и полиомавирус (Б) (электронная микроскопия)
Вирусы гепатита В. Вирус гепатита В включён в состав рода Orthohepadnavirus семейства Hepadnaviridae. Вирионы вируса гепатита В сферической формы 42 нм в диаметре имеют суперкапсид. Геном образует неполная (одна нить короче, рисунок 23) двухнитевая кольцевая молекула ДНК. В состав сердцевины также входит ДНК-зависимая ДНК-полимераза. В динамике процесса репродукции вирусная ДНК интегрирует в ДНК клетки. В крови больных гепатитом В циркулируют частицы трёх морфологических типов. Наиболее часто обнаруживают сферические частицы около 22 нм в диаметре; реже − нитевидной формы около 22 нм в диаметре и 50–230 нм в длину. Вирусные частицы этих типов не проявляют инфекционных свойств. Лишь 7 % частиц представлены комплексными двухслойными сферическими образованиями с полной структурой – частицы Дейна (рисунок 24),проявляющие выраженную инфекционность. Их оболочку на 70 % поверхности образуют белки.
Парвовирусы. Parvoviridae – семейство мелких вирусов, содержащих однонитевую ДНК. Репликация и сборка вирионов происходит в клеточном ядре. Выделяют 3 рода: Parvovirus (типичный вид – вирус крыс Килхэма), Densovirus и род, включающий ассоциирующиеся с тканями миндалин сателлитные вирусы.
|
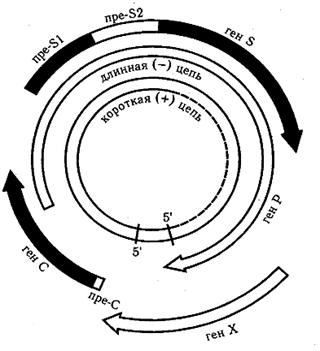
Рисунок 23 – Схема строения генома вируса гепатита В
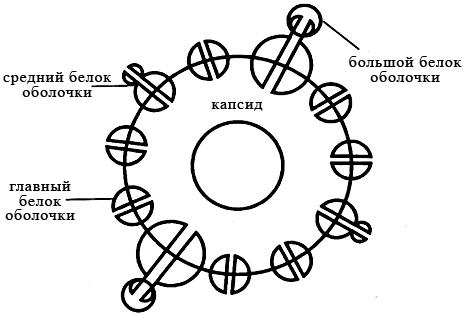
Рисунок 24 – Схема строения частицы Дейна
Тогавирусы. Краснуха – острая инфекция, проявляющаяся кратковременной лихорадкой, мелкопятнистым покраснением кожи, увеличением лимфатических узлов и поражением плода у беременных. Вирус краснухи включён в род Rubivirus семейства Togaviridae и является единственным тогавирусом, эпидемиология которого не связана с членистоногими-переносчиками. Зрелые вирионы имеют сферическую форму и диаметр 50–60 нм. Геном образован несегментированной молекулой +РНК. Липидная оболочка суперкапсида содержит гликопротеины, имеющие форму шипов.
Коронавирусы. Семейство Coronaviridae включает один род – Coronavirus, объединяющий вирионы округлой или овальной формы диаметром 50–220 нм. Зрелые частицы окружены суперкапсидом, включающим редко расположенные гликопротеиновые шипы, состоящие из тонкой шейки и массивной шаровидной овальной или грушевидной головки, что придаёт им сходство с солнечной короной (рисунок 25).
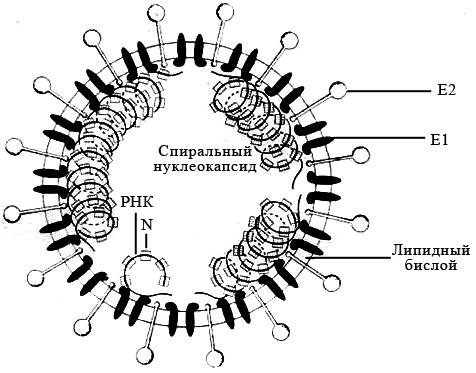
Рисунок 25 – Модель коронавируса
Вирусный нуклеокапсид представляет собой протяженную, несегментированную, гибкую спираль, содержащую геномную +РНК и большое число молекул фосфорилированного нуклеокапсидного белка N. Вирусная оболочка состоит из липидного бислоя, образующегося из внутриклеточной мембраны клетки-хозяина, и двух вирусных гликопротеинов БД и Е2. Матриксный гликопротеин Е1 пронизывает липидный бислой и взаимодействует с нуклеокапсидом внутри вирусной частицы.
У человека коронавирусы вызывают поражения воздухоносных путей (рисунок 26) и желудочно-кишечного тракта.
Филовирусы. Род Filovirus семейства Filoviridae объединяет палочковидные или нитевидные, ветвящиеся вирусы (лат. filum, нить). Средние размеры вирионов 14˟80 нм. Геном образован молекулой РНК. Нуклеокапсид организован по типу спиральной симметрии и образует тяж, покрытый суперкапсидом с гликопротеиновыми шипами длиной 7–10 нм. Для человека патогенны вирусы геморрагических лихорадок Мáрбург и Эбóла. Заболевание регистрируется в экваториальных и субэкваториальных районах Африки.
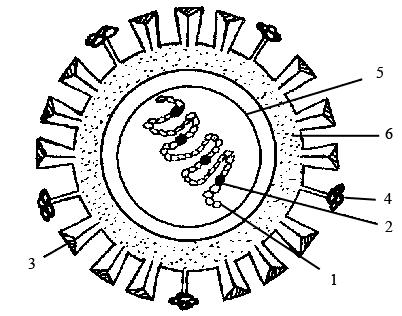
Рисунок 27 – Схема строения вируса гриппа:
1 – спираль рNP; 2 – белки pV1, pV2, pA; 3 – гемагглютинин (500–600 шипов); 4 – нейраминидаза (100–160 шипов); 5 – матриксный белок; 6 – липидный бислой
Репликация ортомиксовирусов первично реализуется в цитоплазме инфицированной клетки; синтез вирусной РНК происходит в ядре.
Наибольшую эпидемическую опасность представляют вирусы гриппа А, вирус гриппа В вызывает локальные вспышки и эпидемии, вирус гриппа С − спорадические случаи гриппа.
Буньявирусы. Семейство Bunyaviridae считается крупнейшим по количеству входящих в него вирусов (около 250). Вирионы буньявирусов имеют сферическую форму и диаметр 90–100 нм. Геном образован молекулой -РНК, состоящей из трех сегментов. Нуклеокапсид организован по типу спиральной симметрии. Снаружи нуклеокапсид покрыт двухслойным липидным суперкапсидом, на котором располагаются белковые структуры с гемагглютинирующей активностью, объединенные в форме поверхностной решетки.
В род Bunyavirus входят возбудители энцефалитов. Вирусы рода Phlebovirus вызывают различные москитные лихорадки. Род Nairovirus включает вирус Конго-крымской геморрагической лихорадки, вызывающей заболевания в России, Молдавии, Украине, на Балканах и в Африке.
Аренавирусы. Характерный морфологический признак представителей семейства Arenaviridae – наличие внутри вирусных частиц электронно-плотных зернистых структур, напоминающих песчаные вкрапления (лат. аrenа, песок). Семейство включает один род Arenavirus, представленный округлыми вирионами диаметром 110–130 нм. Геном образует однонитевая молекула -РНК, содержащая пять сегментов. Вирионы содержат транскриптазу, ответственную за синтез комплементарной нити +РНК, исполняющей роль матрицы. Нуклеокапсид окружен суперкапсидом, на котором расположены многочисленные гликопротеиновые булавовидные шипы. Все аренавирусы относятся к экологической группе робовирусов, и все виды патогенны для человека. Наиболее типичны тяжелые геморрагические лихорадки с высокой летальностью, гриппоподобные поражения, реже серозные минингиты.
Ретровирусы. В состав подсемейства Lentivirinae семейства Retro-viridae входит вирус иммунодефицита человека (ВИЧ). Характерные особенности ретровирусов – уникальное строение генома и наличие обратной транскриптазы (РНК-зависимая ДНК-полимераза). Обратная транскриптаза (или ревертаза) обеспечивает обратную направленность потока генетической информации – не от ДНК к РНК, а наоборот, от РНК к ДНК, в связи с чем семейство и получило своё название (англ., retro, обратно).
Зрелые вирионы ВИЧ имеют сферическую форму, их размеры не превышают 100–120 нм в диаметре.Геном образуют две нити +РНК; их связывают белки р6 и р7 (цифра соответствует молекулярному весу в кД). Капсид образует белок р24. Сердцевина вириона имеет цилиндрическую или конусовидную формы; её формируют белки р18 и р24. В сердцевине располагаются РНК, внутренние белки (р7 и р9), обратная транскриптаза и эндонуклеаза. Матричный белок р17 формирует прослойку между сердцевиной вириона и внешней оболочкой. Суперкапсид образован двойным липидным слоем, который пронизывают гликопротеиновые шипы. Каждый шип состоит из белков gp41 и gpl20 (рисунок 28).
Гликопротеины gp120 локализованы в выступающей части шипа и взаимодействуют с молекулами CD4 на мембранах клеток. Гликопротеины gp41 (белки слияния) располагаются внутри оболочки и обеспечивают её слияние с клеточной мембраной.
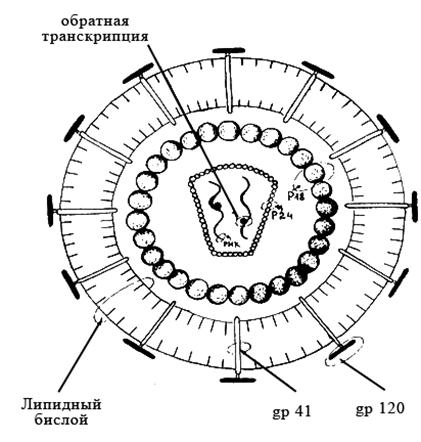
Рисунок 28 – Схема строения вируса иммунодефицита человека
Иридовирусы. Иридовирусы относятся к одноимённому семейству Iridoviridae и роду Iridovirus. В последний включены вирусы радужности насекомых. К семейству Iridoviridae, но к другому предполагаемому роду принадлежит ДНК-содержащий вирус африканской чумы свиней.
Вирусом африканской чумы свиней в природе инфицируются только свиньи (домашние и дикие виды) и мягкие клещи рода Ornithdoros (O. moubata – на юге Сахары в Африке, O. erraticus – в Португалии и Испании). Вирус передается среди клещей трансстадийно, трансовариально и половым путем. Свиньи инфицируются при укусах инфицированными клещами. Болезнь проявляется у домашних и диких европейских свиней.
Иридовирусы пойкилотерных животных выделены только от животных, имеющих водную стадию в цикле своего развития. Большинство иридовирусов насекомых предают синий или бирюзовый цвет инфицированным личинкам.
Вирион икосаэдральной симметрии. Вирусы животных имеют оболочку. Все иридовирусы содержат внутренние липидные мембрано-подобные структуры. Некоторые иридовирусы имеют многочисленные фибры, проходящие через икосаэдрон. Диаметр вирионов 130–170 нм. Это ДНК-геномные вирусы. Молекула ДНК линейная, двуспиральная. Геном иридовирусов позвоночных сильно метилирован.
Собственно вирусы беспозвоночных представлены семейством Baculoviridae, подсемейством Entomopoxvirinae (семейство Poxviridae) и родами Densovirus (семейство Parvoviridae), Iridovirus (семейство Iridoviridae), вирусами насекомых семейства Rhabdoviridae, группой вирусов цитоплазменного полиэдроза (семейство Reoviridae) и группой энтеро-вирусов беспозвоночных семейства Picornaviridae.
Читайте также:
