
Семейство вирусов ана я
Таблица 6.
ОСНОВЫ КЛАССИФИКАЦИИ
ГЛАВА 3. КЛАССИФИКАЦИЯ ВИРУСОВ
КОМПОНЕНТЫ КЛЕТКИ-ХОЗЯИНА
В составе вирионов могут находиться компоненты клетки-хозяина. К таким компонентам могут относиться белки, и даже целые клеточные структуры. Так, например, в составе ряда оболочечных вирусов может находиться белок цитоскелета актин, в составе паповавирусов содержатся клеточные гистоны. Ряд вирусов содержит клеточные ферменты, например протеинкиназы. В составе аренавирусов обнаружены рибосомы.
Клеточные компоненты могут включаться в вирион случайно или закономерно. В некоторых случаях они играют существенную роль в репродукции вируса, как, например, гистоны в репродукции паповавирусов.
Современная классификация вирусов является универсальной для вирусов позвоночных, беспозвоночных, растений и простейших. Она основана на фундаментальных свойствах вирионов, из которых ведущими являются признаки, характеризующие нуклеиновую кислоту, морфологию, стратегию генома и антигенные свойства. Фундаментальные свойства поставлены на первое место, поскольку вирусы со сходными антигенными свойствами обладают и сходным типом нуклеиновой кислоты, сходными морфологическими и биофизическими свойствами. Важным признаком для классификации, который учитывается наряду со структурными признаками, является стратегия вирусного генома, под которой понимают используемый вирусом способ репродукции, обусловленный особенностями его генетического материала. Например, полярность вирусной РНК является основным критерием для группировки вирусов и при отсутствии общих антигенных свойств.
Антигенные и другие биологические свойства являются признаками, лежащими в основе формирования вида и имеющими значение в пределах рода.
В основу современной классификации положены следующие основные критерии.
1. Тип нуклеиновой кислоты (РНК или ДНК), ее структура (количество нитей).
2. Наличие липопротеидной оболочки.
3. Стратегия вирусного генома.
4. Размер и морфология вириона, тип симметрии, число капсомеров.
5. Феномены генетических взаимодействий.
6. Круг восприимчивых хозяев.
7. Патогенность, в том числе патологические изменения в клетках и образование внутриклеточных включений.
8. Географическое распространение.
9. Способ передачи.
10. Антигенные свойства.
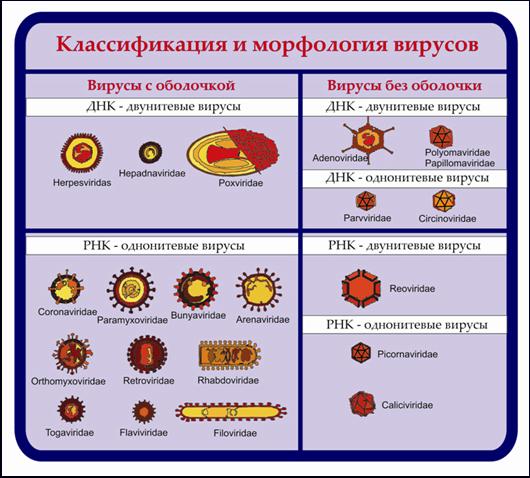 |
Деление на семейства произведено по критериям, изложенным в пунктах 1 и 2, деление на роды и типы — на основании нижеперечисленных признаков. Схематически строение семейств вирионов, поражающих позвоночных, приведено на рис. 6.
Рис. 6. Семейства вирусов позвоночных.
Современная классификация вирусов человека и животных охватывает более 4/5 всех известных вирусов, которые распределены в 19 семейств, из них 7 — ДНК-содержащих и 12 — РНК-содержащих вирусов. Некоторые из этих семейств имеют в своем составе также вирусы беспозвоночных и вирусы растений. К числу семейств вирусов исключительно позвоночных относятся вирусы герпеса, аденовирусы, паповавирусы, гепаднавирусы, парамиксовирусы, ортомиксовирусы, аренавирусы, коронавирусы. Некоторые вирусы обладают уникальной способностью преодолевать филогенетические барьеры и размножаться как в позвоночных, так и в беспозвоночных хозяевах (клещи, комары, москиты). К таким семействам относятся буньявирусы, тогавирусы, роды Vesiculovirus и Lyssavirus семейства рабдовирусов, род Orbivirus семейства реовирусов, вирус африканской лихорадки свиней семейства иридовирусов. Для этих вирусов членистоногие являются и естественными хозяевами, и переносчиками инфекции между позвоночными. Такие вирусы составляют экологическую группу арбовирусов, т. е. вирусов позвоночных, передающихся членистоногими.
Семейства вирусов животных и их таксономические признаки приведены в табл. 6 и 7.
Семейства РНК-содержащих вирусов
• Ортомиксовирусы (вирусы гриппа А, В и С).
• Парамиксовирусы (парагриппозные вирусы, вирус эпидемического паротита,
вирус ньюкастлской болезни, вирус кори, респираторно-синцитиальный вирус).
• Пикорнавирусы более 110 серотипов риновирусов, а также некоторые
серотипы вирусов Коксаки А и В и ЕСНО).
• Реовирусы (3-й серотип).
Семейство ДНК-содержащих вирусов
• Аденовирусы (41 серотип, из них 8 являются возбудителями ОРЗ.
Вирусная инфекция характеризуется накоплением вирусных частиц в клетках организма человека и выходом сформировавшихся вирионов из клеток. Лабораторная диагностика проводится в трёх направлениях:
Выявление внутриклеточных включений, вызванных репродукцией вируса.
Внутриклеточные включения формируются в заражённых вирусом клетках — это места синтеза и сборки вирусных структур. Они являются продуктами взаимосодействия вируса и клетки. Внутриклеточные включения могут быть в цитоплазме и в ядрах зараженных вирусом клетках. По форме включения могут быть округлыми, кристаллоподобными, в виде тяжей и т.п. Для идентификации внутриклеточных включений (принадлежности к тому или другому вирусу) часто используют различные серологические реакции (РИФ, ИФА, РИА, ПЦР, метод молекулярной гибридизации, метод иммуноэлектронной микроскопии и др.).
2. Вирусологическая диагностика, включающая накопление вируса в живых системах (куриный эмбрион, лабораторное животное и различные культуры клеток), индикацию накопленного вируса в живых системах и идентификацию выделенного вируса с помощью серологический реакций, позволяющих установить антигенную структуру выделенного вируса.
Вирусы обладают тропизмом к определённой ткани организма и поэтому в одних клетках репродукция вируса сопровождается гибелью клетки (очаговую мелкозернистую деструкцию вызывают вирусы гриппа, клещевого энцефалита, а гроздевидную дегенерацию - аденовирусы) — цитопатическим действием (ЦПД), а в других клетках вирус репродуцируется, а гибели клеток не наблюдается. Поэтому, в первом случае, индикация вируса проводится по ЦПД, бляшкообразующей активности (способность вируса разрушать клетки под слоем агара или на поверхности хорионаллантоисной оболочки куриного эмбриона) или гибели лабораторного животного. Во втором случае, наличие вируса определяют по выявлению поверхностного гемагглютинирующего антигена вируса. Для этого исследуют аллантоисную жидкость куриного эмбриона, культуральную жидкость (питательная среда для роста клеток, в которые был внесён для накопления вирус) и т.п. Гемагглютинирующий антиген вируса определяют с помощью постановки реакции гемагглютинации (РГА).
Принципиальная схема лабораторной диагностики вирусных и риккетсиозных инфекции
I этап – забор исследуемого материала
1) От больного: носоглоточные смывы, испражнения, спинномозговая жидкость, кровь, отделяемое конъюнктивы глаз, содержимое папул, везикул, пустул.
2) От трупа: кусочки органов и тканей, содержимое кишечника, кровь из полости сердца, спинномозговая жидкость.
3) В очаге: трупы павших животных или кусочки органов и тканей от них, кровососущие насекомые, клеши, вода и т.д.
| Вирусоскопия | Вирусология | Серодиагностика |
| Цель - обнаружение вируса в исследуемом материале. Методы: 1. Риноцитоскопия (выявление внутриклеточных включений – косвенное обнаружение вирусов. По Пигаревскому; по Морозову). 2. Прямая РИФ. 3.Простая люминесцентная микро-скопия (обработка препаратов аурамином позволяет дифференцировать РНК-содержащие (красное свечение) и ДНК-содержащие (зеленое свечение) вирусы) 4. Электронная микроскопия для изучения ультраструктуры вирусов. 5. Прямая РИФ (используют специфические антитела, конъюгированные с ФИТЦ). | Цель — накопление вируса и его идентификация. Исследуемый материал обрабатывается антибиотиками для уничтожения сопутствующей микрофлоры или пропускается через бактериальные фильтры. Для репродукции вирусов существует 3 живые системы (модели). II этап — накопление  Курин.эмбр. Культ. клеток. Лаб. жив III этап – обнаружение вируса (индикация) РГА или ЦПД Клин. проявл. гибель или гибель I V этап - идентификация (сероидентификация) с помощью диагностических иммунных сывороток РТГА, РСК РН, РП, ИФА, и т. д. | Цель – изучение парных сывороток больного с целью выявления динамики нарастания титра специфических Ig (за положительный титр принимается 4-хкратное нарастание титра антител).
|
Если вирус не разрушает клетки, но репродукция его происходит, то репродукцию вируса определяют по феномену гемадсорбции — клетки, продуцирующие вирус, адсорбируют на своей поверхности эритроциты крови различных лабораторных животных. Идентификацию выделенного вируса проводят на основании выявления антигенной структуры выделенного вируса. Для этого используют антитела к различным антигенам вируса и ставят серологические реакции: РСК, РТГА, РН (реакция нейтрализации), РП, РПНА (реакция подавления нейраминидазной активности вируса), ИФА, РИА и др.
3. Серодиагностика — постановка диагноза заболевания на основании иммунобиологической перестройки в организме. Для этого используются различные серологические реакции, позволяющие выявить в крови больных людей или реконвалесцентов циркуляцию различных классов иммуноглобулинов к определённым антигенам вирусной частицы.
При проведении серодиагностики определяют суммарные иммуноглобулины. В этом случае кровь больного (реконвалесцента) исследуют в динамике с интервалом 10-14 дней. Для постановки диагноза заболевания необходимо, чтобы концентрация антител в крови возросла не менее, чем в 4 раза. Кроме этого в крови определяют различные классы иммуноглобулинов к антигенам вирусных частиц. В этом случае используют обычно однократное исследование сыворотки крови больного.
Серодиагностика при гриппе имеет ретроспективное значение и предназначена для подтверждения результатов вирусологических исследований с целью решения вопроса о выборе вакцинного препарата для иммунизации угрожаемых контингентов населения. Определяют подтип вируса гриппа, вызвавшего эпидемию или пандемию.
Достоверное нарастание титров антител к вирусам гриппа отмечается на 2-3 неделе после перенесённого заболевания. В РСК парные сыворотки исследуют одновременно с вирусами трех типов. РСК более чувствительна, чем РТГА, и позволяет диагностировать грипп в очагах до 90% случаев.
РТГА применяется для определения подтипа штамма вируса гриппа, тип которого был определен в РСК. РН применяют в основном для установления штаммовой принадлежности вируса гриппа.
В последнее время для серодиагностики гриппа и др. вирусных инфекций применяют метод непрямой иммунофлюоресценции, позволяющий выявить и титровать специфические противовирусные антитела в сыворотках людей и ставить диагноз в 75% случаев.
РПНА (реакция подавления нейраминидазной активности) используется для идентификации новых штаммов вирусов гриппа. Для изучения антигенной структуры вирусов гриппа используют реакцию иммуноадсорбции, метод радиоиммунного анализа и иммуноэнзимный метод.
Для диагностики парагриппозной инфекции ставят РТГА с 4 антигенами (типовыми) парагриппозных вирусов. В реакции используют эритроциты морских свинок.
Серодиагностика аденовирусных заболеваний проводится на основании нарастания титра антител в парных сыворотках больных и реконвалесцентов. Применяется реакция нейтрализации вируса в культуре клеток, РСК, РТГА. Комплементсвязывающие антитела появляются с 10-15 дня болезни, носят группоспецифический характер и сохраняются несколько лет. Эту реакцию сложно использовать для выявления свежих случаев заболевания, т.к. многие люди имеют постоянно высокие титры антител к аденовирусам. Это связано с тем, что все аденовирусы имеют единый комплементсвязывающий антиген. РН и РТГА ставят с эталонными типами аденовирусов. Четырехкратное нарастание типоспецифических антител свидетельствует об установлении типа аденовируса.
Дата добавления: 2018-10-27 ; просмотров: 303 ;

Вирусы — это микроскопические патогены, заражающие клетки живых организмов для самовоспроизводства. Они состоят из одного вида нуклеиновой кислоты (или ДНК или РНК, но не обе вместе), которая защищена оболочкой, содержащей белки, липиды, углеводы или их комбинацию. Размер типичного вируса варьируется от 15 до 350 нм, поэтому его можно увидеть только с помощью электронного микроскопа.
В 1892 году русский ученый Д.И. Ивановский впервые доказал существование ранее неизвестного типа возбудителя болезней, это был вирус мозаичной болезни табака. А в 1898 году Фридрих Лоффлер и Пол Фрош нашли доказательства того, что причиной ящура у домашнего скота была инфекционная частица, которая меньше, чем любая бактерия. Это были первые шаги к изучению природы вирусов, генетических образований, которые лежат где-то в серой зоне между живыми и неживыми состояниями материи. На текущий момент описано около 6 тыс. вирусов, но их существует несколько миллионов.
Строение вирусов
Вне клеток-хозяев вирусы существуют в виде белковой оболочки (капсида), иногда заключенного в белково-липидную мембрану. Капсид обволакивает собой либо ДНК, либо РНК, которая кодирует элементы вируса. Находясь в такой форме вне клетки, вирус метаболически инертен и называется вирионом.
Простая структура, отсутствие органелл и собственного метаболизма позволяет некоторым вирусам кристаллизоваться, т.е. они могут вести себя подобно химическим веществам. С появлением электронных микроскопов было установлено, что их кристаллы состоят из тесно прижатых друг к другу нескольких сотен миллиардов частиц. В одном кристалле вируса полиомиелита столько частиц, что ими можно заразить не по одному разу всех жителей Земли.
Формы вирусов
Вирусы встречаются в трех основных формах. Они бывают:
- Сферическими (кубическими или полигидральными). Вирусы герпеса, типулы, полиомы и т.д.
- Спиральными (цилиндрическими или стержнеобразными). Вирусы табачной мозаики, гриппа, эпидемического паротита и др.
- Сложными. Например, бактериофаги.
Проникновение вирусов в клетку-хозяина
Капсид в основном защищает нуклеиновую кислоту от действия клеточного нуклеазного фермента. Но некоторые белки капсида способствуют связыванию вируса с поверхностью клеток-хозяев, и работают, как ключики, вставляемые в нужные замочки. Другие поверхностные белки действуют как ферменты, они растворяют поверхностный слой клетки-хозяина и таким образом помогают проникновению нуклеиновой кислоты вируса в клетку-хозяина.
Жизненный цикл вирусов сильно отличается у разных видов, но существует шесть основных этапов жизненного цикла вирусов:
Присоединение к клетке-хозяину представляет собой специфическое связывание между вирусными капсидными белками и рецепторами на клеточной поверхности. Эта специфика определяет хозяина вируса.
Проникновение следует за прикреплением: вирионы проникают в клетку-хозяина через рецептор-опосредованный эндоцитоз или слияние мембран. Это часто называют вирусной записью.
Проникновение вирусов в клетку достигается за счет:
Размножение вирусов
После того, как вирусный геном освобождается от капсида, начинается его транскрипция или трансляция. Именно эта стадия вирусной репликации сильно различается между ДНК- и РНК-вирусами и вирусами с противоположной полярностью нуклеиновой кислоты. Этот процесс завершается синтезом новых вирусных белков и генома (точных копий внедрённых).
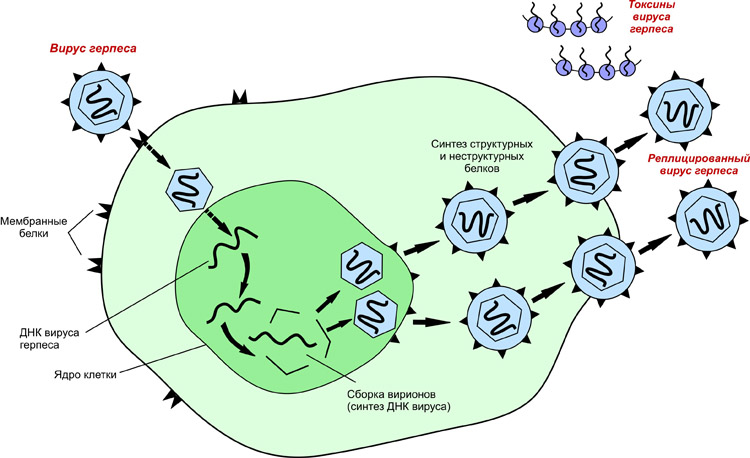
Механизм репликации зависит от вирусного генома.
- ДНК-вирусы обычно используют белки и ферменты клетки-хозяина для получения дополнительной ДНК, она транскрибируется в РНК-мессенджер (мРНК), которая затем используется для управления синтезом белка.
- РНК-вирусы обычно используют ядро РНК в качестве матрицы для синтеза вирусной геномной РНК и мРНК. Вирусная мРНК направляет клетку-хозяина на синтез вирусных ферментов и капсидных белков и сборку новых вирионов. Конечно, есть исключения из этого шаблона. Если клетка-хозяин не обеспечивает ферменты, необходимые для репликации вируса, вирусные гены предоставляют информацию для прямого синтеза отсутствующих белков.
Чтобы преобразовать РНК в ДНК, вирусы должны содержать гены, которые кодируют вирус-специфический фермент обратной транскриптазы. Она транскрибирует матрицу РНК в ДНК. Обратная транскрипция никогда не происходит в неинфицированных клетках. Необходимый фермент, обратная транскриптаза, происходит только от экспрессии вирусных генов в инфицированных клетках.
Вироиды
Вироиды заражают только растения. Одни вызывают экономически важные заболевания сельскохозяйственных культур, в то время как другие являются доброкачественными. Двумя примерами экономически важных вироидов являются кокосный cadang-cadang (он вызывает массовую гибель кокосовых пальм) и вироид рубцовой кожицы яблок, который безнадежно портит товарный вид яблок.
30 известных вироидов были классифицированы в две семьи.
- Члены семейства Pospiviroidae, названные по имени вироида клубневого веретена картофеля, имеют палочковидную вторичную структуру с небольшими одноцепочечными областями, имеет центральную консервативную область, и реплицируются в ядре клетки.
- Avsunviroidae, названный в честь вироида авокадо, имеет как палочковидную, так и разветвленную области, но не имеет центральной консервативной области и реплицируется в хлоропластах растительной клетки.
В отличие от вирусов, которые являются паразитами механизма трансляции хозяина, вироиды являются паразитами клеточных транскрипционных белков.
Бактериофаги

Существуют тысячи разновидностей фагов, каждый из которых может заразить только один тип или несколько близких типов бактерий или архей. Фаги классифицируются по ряду семейств вирусов; например:
Как и все вирусы, фаги являются простыми организмами, которые состоят из ядра генетического материала (нуклеиновой кислоты), окруженного капсидом белка. Нуклеиновая кислота может представлять собой либо ДНК, либо РНК, и может быть двухцепочечной или одноцепочечной.
Существует три основных структурных формы фага:
- Икосаэдрическая (20-сторонняя) головка с хвостом
- Икосаэдрическая головка без хвоста
- Нитевидная форма
Во время заражения фаг прикрепляется к бактерии и вставляет в нее свой генетический материал. После этого фаг обычно следует одному из двух жизненных циклов: литическому (вирулентному) или лизогенному (умеренному).
Литические, или вирулентные, фаги захватывают механизм клетки, чтобы скопировать компоненты фага. Затем они разрушают или лизируют клетку, высвобождая новые частицы фага.
Лизогенные, или умеренные, фаги включают свою нуклеиновую кислоту в хромосому клетки-хозяина и реплицируются с ней как единое целое, не разрушая клетку. При определенных условиях лизогенные фаги могут индуцироваться в соответствии с литическим циклом.
Существуют и другие жизненные циклы, в т.ч. псевдолизогенез и хроническая инфекция. При псевдолизогении бактериофаг проникает в клетку, но не использует механизм репликации клеток и не интегрируется в геном хозяина, просто как бы прячется внутри бактерии, не нанося ей никакого вреда. Псевдолизогенез возникает, когда клетка-хозяин сталкивается с неблагоприятными условиями роста и, по-видимому, играет важную роль в выживании фага, обеспечивая сохранение генома фага до тех пор, пока условия роста хозяина снова не станут благоприятными.
При хронической инфекции новые фаговые частицы образуются непрерывно и длительно, но без явного уничтожения клеток.
Вскоре после открытия фаги начали использовать для лечения бактериальных заболеваний человека, таких как бубонная чума и холера. Но фаговая терапия тогда не была успешной, и после открытия антибиотиков в 1940-х годах она была практически заброшена. Однако с появлением устойчивых к антибиотикам бактерий терапевтическому потенциалу фагов уделяется все больше внимания.
Наше время с антибиотиками заканчивается. В 2016 году женщина в штате Невада умерла от бактериальной инфекции, вызванной Klebsiella pneumoniae, которая была устойчивой ко всем известным антибиотикам. Бактерии, устойчивые к колистину, антибиотику последней инстанции, были обнаружены на свинофермах в Китае. В настоящее время бактерии приспосабливаются к антибиотикам быстрее, чем когда-либо.
Покажите ножницы которыми вирусы разрезают молекулу РНК что бы встроиться для мутации.Может что нибудь придумаете другое.К примеру деление цепочка аминокислот получив энергию из вне как одноименные заряды распадается на две. К каждой соединятся только те какие были ранее (другие проскочат мимо),казалось бы копии,но внутренняя энергия разная(уменьшается увеличивается) поэтому распад и создание. Вся химия углерода на этом построена 1000 орган соединений создает у других хим элементов этого свойства нет. Иммунная система делает накладку(интерференция)с помощью энергии интерферонов пытаясь разрушить цепочку РНК вируса.Надо помочь организму но не вакциной(вирус быстро мутирует)
Алексей! Я полный профан в этой области! И если вы сможете, ответьте мне на вопрос: вирусы состоят из молекул?Ещё раз говорю: если с Вашей точки зрения мой вопрос дурацкий, посмейтесь и,если не трудно,ответьте доступным языком. Спасибо!
Среди онкогенных вирусов существует две большие группы – группа эндогенных онковирусов, способных передаваться вертикально (от отца к потомкам) и группа экзогенных онковирусов, распространяющихся горизонтально (среди одного поколения) по типу обычных возбудителей инфекции. Переходной группой являются онкогенные эндогенные вирусы, которые могут распространяться горизонтально и заражать близкие биологические виды.
Вирусы, индуцирующие злокачественные образования, можно разделить на две группы с различными физическими, химическими и биологическими свойствами.
Онкогенные вирусы, как и все другие вирусы, подразделяются на ДНК- содержащие и РНК- содержащие, представленные определенным количеством семейств.
Семейство Papovaviridae
Род Papillomavirus (вирусы папилломы людей, кроликов, собак, коров).
Род Polyomavirus (вирусы полиомы мышей, вакуолизирующий вирус SV40 обезьян, вирусы многоочагованной лейкоэнцефалопатии людей).
Семейство Adenoviridae
Типы: 3, 7, 11, 12, 14, 16, 18, 21, 31.
Семейство Herpviridae
Род Herpesvirus (тип 2, вирус лимфомы кроликов, обезьян, вирус EВ, вызывающий у людей инфекционный мононуклеоз)
Семейство Poxviridae
Род Lepovirus (вирус контагиозного моллюска людей).
Семейство Parvoviridae
Род Parvovirus (вирусы панлейкимии крыс и некоторые латентные вирусы)
Семейство Hepadnaviridae (вирус гепатоцеллюлярной аденокарциномы, вирус гепатита В (сывороточного)
Семейство Flavoviridae (вирус гепатита ни А ни В)
Семейство Retroviridae
Подсемейство Onkovirinae – вирусы групп B,C,D, вирус бычьего лейкоза, вирусы клеточной лейкимии человека (HTLV-I, HTLV-II), вирус лимфолейкоза человека.
Подсемейство Lentivirinae – HIV -1, HIV- 2.
Подсемейство Spumavirinae – вирус лейкоза мышей.
Многие из исследователей считают, что в большинстве случаев образование злокачественных опухолей обуславливается РНК- содержащими онкогенными вирусами.
Онкогенные вирусы, содержащие РНК, выделены в самостоятельное семейство Retroviridae, в состав которого входят более 100 видов. Основные представители отнесены к подсемейству онковирусов, в состав которого входят онковирусы типов B,C,D и бычьего лейкоза.
Наиболее распространены онковирусы типа С. Их обнаруживают у рыб, птиц, рептилий, млекопитающих, в том числе и у людей.
Вирусы типа В обнаружены у мышей и морских свинок, вирусы типа D – у обезьян.
Онковирусы типа D были выделены из перевиваемых клеток человека. Считают, что исходными клетками, в которых репродуцируется этот вирус, являются клетки HeLa (эти клетки были выделены из злокачественной опухоли женщины, погибщей от рака, Гелены Лайне).
По морфологии РНК- содержащие онковирусы имеют сферическую форму диаметром 80 – 100 нм. В середине вириона находится нуклеоид, окруженный белковой мембраной, поверх которой располагается глюцидо-липидо-протеиновая оболочка.
РНК- онковирусы имеют сложный химический состав. Зрелые вирионы имеют содержат 2% РНК, 60% белка, 35% липидов, 3,5% углеводов.
Среди белков онковирусов особое место принадлежит ферментам, часть которых имеет клеточное происхождение и попадает в вирион при его формировании. К таким ферментам относятся РНК- и ДНК- зависимая ДНК-полимераза, ДНК- экзонуклеаза, РНК- аза, лигаза, протеинкиназа и др. (свыше 10 наименований).
Наиболее характерным ферментом онковирусов является обратная транскриптаза, и ее наличие в клетке может служить одним из тестов инфицирования тканевых клеток онкогенным вирусом.
Онковирусы имеют очень сложную антигенную структуру. Группоспецифические антигены связаны с нуклеопротеидом, окруженным белковой оболочкой. Их выявляют с помощью иммунодифузии, иммунной флюоресценции, РСК.
Типоспецифические антигены связаны с гикопротеидами и выявляются с помощью иммунодифузии, иммунной флюоресценции, РСК и РН.
Все онкогенные РНК- содержащие вирусы чувствительны к эфиру, формалину, кислой среды (рН 4,5). Легко инактивируются при 56 градусах на протяжении 30 минут, сравнительно устойчивы к ультрафиолетовым лучам и хорошо переносят минусовые температуры.
Онкогенные ДНК- содержащие вирусы дифференцируются друг от друга по величине, морфологии вириона, антигенной структуре, содержанием генетического.
Так, для паповавирусов, аденовирусов и парвавирусов общим является кубический тип симметрии и отсутствие внешней оболочки. В силу того, что вирусы не содержат липиды, они устойчивы к эфиру, хлороформу, слабых кислот. Хорошо переносят действие низких температур и высушивание, но чувствительны к нагреванию и при 56 градусах погибают через 10 – 15 минут.
Репродуцируются паповавирусы и аденовирусы в ядре клетки хозяина.
Клетки пораженные паповавирусами и аденовирусами содержат некоторые вирусиндуцированные антигены, специфические для трансформирующего вируса. К ним относятся Т-антигены опухолей. По химической структуре Т-антиген относится к фоспротеину. Т-антиген формируется в ядре на ранних этапах репликационного цикла перед началом синтеза вирусной ДНК и белков вирусного капсида.
Выявление этого антигена проводится методами иммунофлюоресценции или РСК при помощи антител, накапливающихся в сыворотке крови животных, у которых имеются первичные или транплантационые опухоли.
К другой группе антигенов, которые обнаруживаются в индуцированных паповавирусами и аденовирусами клетках, относятся опухолевые специфические трансплантационные антигены, которые удается выявить из – за их способности блокировать вирусный канзерогенез.
В последнее время вирусам герпеса, как потенциальным онкогенным агентам, уделяется все большее внимание.
Вирус простого герпеса типа 2 (ВПГ -2) у людей передается половым путем, Серологические и эпидемиологические исследования выявили связь между заражением этим вирусом и инвазиозным раком шейки матки. Изучение клеток, трансформированных ВПГ-2, показало, что в них находится не весь геном вируса, а только небольшие фрагменты вирусной ДНК.
Вирус герпеса ЕВ (Эпштейна – Барра) вызывает широко распространенный среди людей инфекционный мононуклеоз. ДНК вируса ЕВ и антигены вируса ЕВ обнаруживаются при двух злокачественных опухолях – лимфоме Беркита (поражение верхней челюсти) и карциноме носоглотки.
Серологические и эпидемиологические исследования показали, что инфицирование вирусом ЕВ распространено повсеместно.
Вирус обладает тропизмом к лимфоидным клеткам, в частности, к В- лимфоцитам человека.
Антитела по отношению к антигенам вируса ЕВ у больных выявляются при помощи иммунофлюоресценции, РСК и диффузии в геле.
Иммунологические исследования, проведенные у больных раком предстательной железы, позволили установить, что трансформация нормальных клеток в злокачественные обуславливается вирусом цитомегалии.
К онкогенным вирусам относится вирус гепатита В, трансформирующий клетки печени, в которые он проникает. У больных первичной злокачественной опухолью печени HbsAg встречается в сыворотке крови в 95% случаев.
К онкогенным вирусам относится также вирус гепатита С (один из вирусов возбудителей гепатита «ни А ни В).
Классификация вирусов сложна и в настоящее время остается незавершённой. Классификационные принципы К. Линнея в отношении вирусов оказались неприемлемыми. С 1971 г. во всем мире официально используемой является Универсальная система классификации вирусов, основанная на политетическом иерархическом принципе, выдвинутом М. Адансоном (1727-1806 гг.) - французским ботаником, одним из основоположников естественной системы растений и применения математических методов в биологии. По этому принципу учитывается максимальное число морфологических, физиологических, генетических и прочих признаков вирусов (не менее 60), каждый из которых имеет определенную степень значимости. Недостатком адансоновой классификации является не её трудоёмкость, а отсутствие полной информации о многих вирусах.
Согласно последней Универсальной классификации вирусов, принятой Международным Комитетом по таксономии вирусов (МКТВ) в 1998 г. (Сан-Диего, США), выделяется 3 порядка, 56 семейств, 9 подсемейств, 203 рода, 30 неопределённых родов, а также 3954 вида, штаммов и/или серотипов видов и неопределённых видов вирусов. Вирусные таксоны выше порядков, например, подкласс, класс, подцарство, царство, в классификации вирусов не учреждены.
Порядки вирусов – это группы семейств вирусов с общими характеристиками, отличающиеся от других порядков и семейств. Обозначаются названиями с суффиксами -virales. В настоящее время выделено 3 порядка вирусов: Mononegavirales (вирусы, содержащие однониточные негативные РНК), Caudovirales (“хвостатые” бактериофаги), Nidovirales (от лат. nido – гнездо; вирусы, у которых субгеномный РНК-сплайсинг транскрибируется во время репликации в форме гнезда), из которых утвержденным МКТВ является только первый.
Порядок Mononegavirales включает семейства: Paramyxoviridae, Rhabdoviridae и Filoviridae.
Семейства вирусов – это группы родов вирусов с общими характеристиками, отличающиеся от вирусов других семейств. Обозначаются названием с суффиксом -viridae. Семейство – термин, который выдержал проверку временем и не изменяется. Этот уровень таксономической иерархии является стабильным краеугольным камнем. Многие семейства вирусов имеют четкое отличие строения вириона, структуру генома, и способ репликации, определяющие филогенетическую независимость или большую филогенетическую отдельную структуру.
Подсемейства выделены в 4 семействах: Poxviridae, Herpesviridae, Parvoviridae, Paramyxoviridae – для более сложной иерархии таксона. Обозначаются суффиксом -virinae.
Критерии, используемые для описания семейства вирусов согласно системе классификации М. Адансона (взяты из указаний МКТВ для описания семейств):
Свойства вириона. 1. Морфология вириона (размер, форма, наличие или отсутствие суперкапсида и пепломеров, симметрия и структура капсида). 2. Физические свойства вириона (молекулярная масса, плотность, коэффициент седиментации, рН-стабильность, температурная устойчивость, катионная (магний и марганец) стабильность, устойчивость к растворителям, детергентам и действию лучистой энергии). 3. Свойства генома (тип нуклеиновой кислоты, количество нитей, линейный или циркулярный, полярность, количество сегментов, размер генома или его сегментов, наличие или отсутствие кэпа на 5-конце, наличие или отсутствие на 5-конце ковалентно связанного полипептида, наличие или отсутствие на 3-конце поли(А)последовательности или другой специфической последовательности, последовательность нуклеотидов). 4. Свойства белков (количество, размеры, функциональная активность, последовательность аминокислот). 5. Липиды (наличие или отсутствие, природа). 6. Карбогидраты (присутствие или отсутствие, природа).
Организация генома и репликация. 1. Организация генома. 2. Способ репликации нуклеиновых кислот. 3. Характеристики транскрипции. 4. Характеристики трансляции и посттрансляционного процессинга. 5. Место аккумуляции белков вириона, место сборки, место высвобождения. 6. Цитопатология.
Антигенные свойства. 1. Серологические особенности. 2. Эпитопы.
Биологические свойства. 1. Круг хозяев, природных и экспериментальных. 2. Патогенность, связь с заболеваниями. 3. Тканевой тропизм, патология, гистопатология. 4. Пути передачи в природе. 5. Векторные взаимоотношения. 6. Географическая распространенность.
Рода вирусов – группы видов с общими характеристиками, отличающиеся от вирусов других родов. Обозначаются терминами с суффиксом -virus. Этот иерархический уровень также является стабильным. Критерии для создания родов различаются в разных семействах: используются все меньшие генетические, структурные и другие особенности для создания новых родов. Основой является общее филогенетическое происхождение (как и для семейств), но это не постулат для будущего.
Вид вирусов – это политетический класс вирусов, имеющий свою тактику репликации и занимающий особую экологическую нишу. Это определение утверждено МКТВ в 1991 г. Члены политетического класса отличаются более чем одним свойством друг от друга, и ни одно из свойств не является решающим. Большим преимуществом этого определения является то, что оно учитывает природную (врожденную) вариабельность вирусов и не зависит от существования одной уникальной характеристики.
Примеры названия видов:
1. Порядок Mononegavirales, семейство Rhabdoviridae, род Lyssavirus, вид Rhabdovirus.
2. Семейство Herpesviridae, подсемейство Alphaherpesvirinae, род Simplexvirus, вид – простого герпеса вирус 2.
Классификация вирусов, патогенных для человека, представлена в табл. 1.
Классификация вирусов, патогенных для человека
Читайте также:
