
Репликация и сборка вируса
Лекция 6
РЕПЛИКАЦИЯ
Репликация ДНК осуществляется ДНК-полимеразами. Для начала репликации необходим предварительный синтез короткого участка РНК на матрице ДНК, который называется затравкой. С затравки начинается синтез нити ДНК, после чего РНК быстро удаляется с растущего участка.
Репликация вирусных ДНК. Репликация генома ДНК-содержащих вирусов в основном катализируется клеточными фрагментами и механизм ее сходен с механизмом репликации клеточной ДНК.
Каждая вновь синтезированная молекула ДНК состоит из одной родительской и одной вновь синтезированной нити. Такой механизм репликации называется полуконсервативным.
У вирусов, содержащих кольцевые двунитчатые ДНК (паповавирусы), разрезается одна из нитей ДНК, что ведёт к раскручиванию и снятию супервитков на определенном участке молекулы.
При репликации однонитчатых ДНК (семейство парвовирусов) происходит образование двунитчатых форм, которые представляют собой промежуточные репликативные формы.
Репликация вирусных РНК. В клетке нет ферментов, способных осуществить репликацию РНК. Поэтому ферменты, участвующие в репликации, всегда вирусспецифические. Репликацию осуществляет тот же фермент, что и транскрипцию; репликаза является либо модифицированной транскриптазой, либо при репликации соответствующим образом модифицируется матрица.
Репликативные комплексы ассоциированы с клеточными структурами либо с предсуществующими, либо вирусиндуцируемыми. Например, репликативные комплексы пикорнавирусов ассоциированы с мембранами эндоплазматической сети, вирусов оспы — с цитоплазматическим матриксом, репликативные комплексы аденовирусов и вирусов герпеса в ядрах находятся в ассоциации со вновь сформированными волокнистыми структурами и связаны с ядерными мембранами. В зараженных клетках может происходить усиленная пролиферация клеточных структур, с которыми связаны репликативные комплексы, или их формирование из предсуществующего материала. Например, в клетках, зараженных пикорнавирусами, происходит пролиферация гладких мембран. В клетках, зараженных реовирусами, наблюдается скопление микротрубочек; в клетках, зараженных вирусами оспы, происходит формирование цитоплазматического матрикса.
В репликативных комплексах одновременно с синтезом геномных молекул осуществляется транскрипция и происходит сборка нуклеокапсидов и сердцевин, а при некоторых инфекциях — и вирусных частиц. О сложной структуре репликативных комплексов говорит, например, такой состав репликативного комплекса аденовирусов: реплицирующиеся ДНК, однонитчатые ДНК, однонитчатые РНК, ферменты репликации и транскрипции, структурные и неструктурные вирусные белки и ряд клеточных белков.
СБОРКА ВИРУСНЫХ ЧАСТИЦ
Синтез компонентов вирусных частиц в клетке разобщен и может протекать в разных структурах ядра и цитоплазмы. Вирусы, репликация которых проходит в ядрах, условно называют ядерными. В основном это ДНК-содержащие вирусы: аденовирусы, паповавирусы, парвовирусы, вирусы герпеса. Вирусы, реплицирующиеся в цитоплазме, называют цитоплазматическими. К ним относятся из ДНК-содержащих вирус оспы и большинство РНК-содержащих вирусов, за исключением ортомиксовирусов и ретровирусов. Однако это разделение весьма относительно, потому что в репродукции тех и других вирусов есть стадии, протекающие соответственно в цитоплазме и ядре.
Внутри ядра и цитоплазмы синтез вирусспецифических молекул также может быть разобщен. Так, например, синтез одних белков осуществляется на свободных полисомах, а других — на полисомах, связанных с мембранами. Вирусные нуклеиновые кислоты синтезируются в ассоциации с клеточными структурами вдали от полисом, которые синтезируют вирусные белки. При таком дисъюнктивном способе репродукции образование вирусной частицы возможно лишь в том случае, если вирусные нуклеиновые кислоты и белки обладают способностью при достаточной концентрации узнавать друг друга в многообразии клеточных белков и нуклеиновых кислот и самопроизвольно соединяться друг с другом, т. е. способны к самосборке.
В основе самосборки лежит специфическое белок-нуклеиновое и белок-белковое узнавание, которое может происходить в результате гидрофобных, солевых и водородных связей, а также стерического соответствия. Белок-нуклеиновое узнавание ограничено небольшим участком молекулы нуклеиновой кислоты и определяется уникальными последовательностями нуклеотидов в некодирующей части вирусного генома. С этого узнавания участка генома вирусными капсидными белками начинается процесс сборки вирусной частицы. Присоединение остальных белковых молекул осуществляется за счет специфичеческих белокбелковых взаимодействий или неспецифических белокнуклеиновых взаимодействий.
В связи с разнообразием структуры вирусов животных разнообразны и способы формирования вирионов, однако можно сформулировать следующие общие принципы сборки.
1. У просто устроенных вирусов формируются провирионы, которые затем в результате модификаций белков превращаются в вирионы. У сложно устроенных вирусов сборка осуществляется многоступенчато. Сначала формируются нуклеокапсиды или сердцевины, с которыми взаимодействуют белки наружных оболочек.
2. Сборка сложно устроенных вирусов (за исключением сборки вирусов оспы и реовирусов) осуществляется на клеточных мембранах. Сборка ядерных вирусов происходит с участием ядерных мембран, сборка цитоплазматических вирусов — с участием мембран эндоплазматической сети или плазматической мембраны, куда независимо друг от друга прибывают все компоненты вирусной частицы.
5. Сложно устроенные вирусы для построения своих частиц используют ряд элементов клетки-хозяина, например липиды, некоторые ферменты, у ДНК-геномного SV40 — гистоны, у оболочечных РНК-геномных вирусов — актин, а в составе ареновирусов обнаружены даже рибосомы. Клеточные молекулы несут определенные функции в вирусной частице, однако включение их в вирион может явиться и следствием случайной контаминации, как, например, включение ряда ферментов клеточных оболочек или клеточных нуклеиновых кислот.
Сборка РНК-содержащих вирусов. Сборка просто устроенных РНК-содержащих вирусов заключается в ассоциации вирусного генома с вирусными капсидными белками с образованием нуклеокапсида.
Все вирусные компоненты — нуклеокапсиды и суперкапсидные белки прибывают к месту сборки независимо друг от друга. Первыми к месту сборки прибывают суперкапсидные белки. Обычно этими белками являются гликопротеиды, которые синтезируются в полисомах, связанных с мембранами, и через шероховатые, а затем гладкие мембраны в результате слияния с ними везикул комплекса Гольджи транспортируются на наружную поверхность плазматических мембран или остаются в составе везикул.
Включение М-белка в клеточные мембраны является сигналом для сборки вирусной частицы: вслед за включением немедленно следует связывание нуклеокапсидов с мембранами и почкование вирусной частицы. Тем самым М-белок обладает функцией лимитирующего сборку фактора.
В результате связывания ДНК с капсидами появляется новый класс промежуточных форм, которые называются неполными формами. Помимо неполных форм с разным содержанием ДНК, существует другая промежуточная форма в морфогенезе — незрелые вирионы, отличающиеся от зрелых тем, что содержат ненарезанные предшественники полипептидов. Таким образом, морфогенез вирусов тесно связан с модификацией (процессингом) белков.
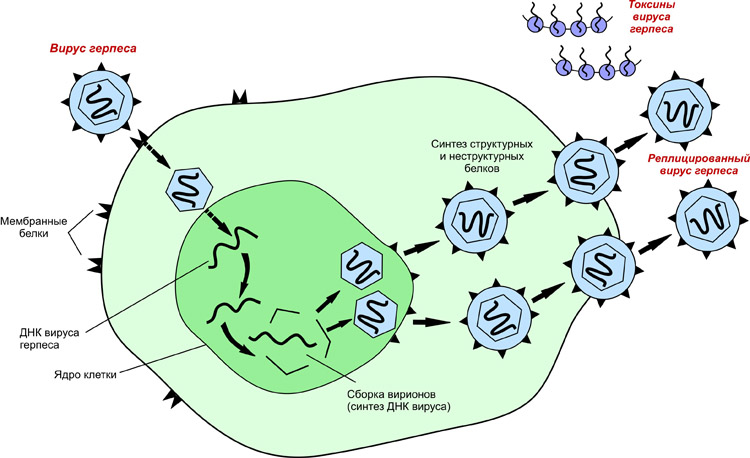
Сборка ядерных вирусов начинается в ядре, обычно — с ассоциации с ядерной мембраной. Формирующиеся в ядре промежуточные формы вируса герпеса почкуются в перинуклеарное пространство через внутреннюю ядерную мембрану, и вирус приобретает таким путем оболочку, которая является дериватом ядерной мембраны. Дальнейшая достройка и созревание вирионов происходит в мембранах эндоплазматической сети и в аппарате Гольджи, откуда вирус в составе цитоплазматических везикул транспортируется на клеточную поверхность.
ВЫХОД ВИРУСНЫХ ЧАСТИЦ ИЗ КЛЕТКИ
Выход из клетки путем взрыва связан с деструкцией клетки, нарушением ее целостности, в результате чего находящиеся внутри клетки зрелые вирусные частицы оказываются в окружающей среде. Такой способ выхода из клетки присущ вирусам, не содержащим липопротеидной оболочки (пикорна-, рео-, парво-, папова-, аденовирусы). Однако некоторые из этих вирусов могут транспортироваться на клеточную поверхность до гибели клетки.
Выход из клеток путем почкования присущ вирусам, содержащим липопротеидную мембрану, которая является дериватом клеточных мембран. При этом способе клетка может длительное время сохранять жизнеспособность и продуцировать вирусное потомство, пока не произойдет полное истощение ее ресурсов.

Вирусы — это микроскопические патогены, заражающие клетки живых организмов для самовоспроизводства. Они состоят из одного вида нуклеиновой кислоты (или ДНК или РНК, но не обе вместе), которая защищена оболочкой, содержащей белки, липиды, углеводы или их комбинацию. Размер типичного вируса варьируется от 15 до 350 нм, поэтому его можно увидеть только с помощью электронного микроскопа.
В 1892 году русский ученый Д.И. Ивановский впервые доказал существование ранее неизвестного типа возбудителя болезней, это был вирус мозаичной болезни табака. А в 1898 году Фридрих Лоффлер и Пол Фрош нашли доказательства того, что причиной ящура у домашнего скота была инфекционная частица, которая меньше, чем любая бактерия. Это были первые шаги к изучению природы вирусов, генетических образований, которые лежат где-то в серой зоне между живыми и неживыми состояниями материи. На текущий момент описано около 6 тыс. вирусов, но их существует несколько миллионов.
Строение вирусов
Вне клеток-хозяев вирусы существуют в виде белковой оболочки (капсида), иногда заключенного в белково-липидную мембрану. Капсид обволакивает собой либо ДНК, либо РНК, которая кодирует элементы вируса. Находясь в такой форме вне клетки, вирус метаболически инертен и называется вирионом.
Простая структура, отсутствие органелл и собственного метаболизма позволяет некоторым вирусам кристаллизоваться, т.е. они могут вести себя подобно химическим веществам. С появлением электронных микроскопов было установлено, что их кристаллы состоят из тесно прижатых друг к другу нескольких сотен миллиардов частиц. В одном кристалле вируса полиомиелита столько частиц, что ими можно заразить не по одному разу всех жителей Земли.
Формы вирусов
Вирусы встречаются в трех основных формах. Они бывают:
- Сферическими (кубическими или полигидральными). Вирусы герпеса, типулы, полиомы и т.д.
- Спиральными (цилиндрическими или стержнеобразными). Вирусы табачной мозаики, гриппа, эпидемического паротита и др.
- Сложными. Например, бактериофаги.
Проникновение вирусов в клетку-хозяина
Капсид в основном защищает нуклеиновую кислоту от действия клеточного нуклеазного фермента. Но некоторые белки капсида способствуют связыванию вируса с поверхностью клеток-хозяев, и работают, как ключики, вставляемые в нужные замочки. Другие поверхностные белки действуют как ферменты, они растворяют поверхностный слой клетки-хозяина и таким образом помогают проникновению нуклеиновой кислоты вируса в клетку-хозяина.
Жизненный цикл вирусов сильно отличается у разных видов, но существует шесть основных этапов жизненного цикла вирусов:
Присоединение к клетке-хозяину представляет собой специфическое связывание между вирусными капсидными белками и рецепторами на клеточной поверхности. Эта специфика определяет хозяина вируса.
Проникновение следует за прикреплением: вирионы проникают в клетку-хозяина через рецептор-опосредованный эндоцитоз или слияние мембран. Это часто называют вирусной записью.
Проникновение вирусов в клетку достигается за счет:
Размножение вирусов
После того, как вирусный геном освобождается от капсида, начинается его транскрипция или трансляция. Именно эта стадия вирусной репликации сильно различается между ДНК- и РНК-вирусами и вирусами с противоположной полярностью нуклеиновой кислоты. Этот процесс завершается синтезом новых вирусных белков и генома (точных копий внедрённых).
Механизм репликации зависит от вирусного генома.
- ДНК-вирусы обычно используют белки и ферменты клетки-хозяина для получения дополнительной ДНК, она транскрибируется в РНК-мессенджер (мРНК), которая затем используется для управления синтезом белка.
- РНК-вирусы обычно используют ядро РНК в качестве матрицы для синтеза вирусной геномной РНК и мРНК. Вирусная мРНК направляет клетку-хозяина на синтез вирусных ферментов и капсидных белков и сборку новых вирионов. Конечно, есть исключения из этого шаблона. Если клетка-хозяин не обеспечивает ферменты, необходимые для репликации вируса, вирусные гены предоставляют информацию для прямого синтеза отсутствующих белков.
Чтобы преобразовать РНК в ДНК, вирусы должны содержать гены, которые кодируют вирус-специфический фермент обратной транскриптазы. Она транскрибирует матрицу РНК в ДНК. Обратная транскрипция никогда не происходит в неинфицированных клетках. Необходимый фермент, обратная транскриптаза, происходит только от экспрессии вирусных генов в инфицированных клетках.
Вироиды
Вироиды заражают только растения. Одни вызывают экономически важные заболевания сельскохозяйственных культур, в то время как другие являются доброкачественными. Двумя примерами экономически важных вироидов являются кокосный cadang-cadang (он вызывает массовую гибель кокосовых пальм) и вироид рубцовой кожицы яблок, который безнадежно портит товарный вид яблок.
30 известных вироидов были классифицированы в две семьи.
- Члены семейства Pospiviroidae, названные по имени вироида клубневого веретена картофеля, имеют палочковидную вторичную структуру с небольшими одноцепочечными областями, имеет центральную консервативную область, и реплицируются в ядре клетки.
- Avsunviroidae, названный в честь вироида авокадо, имеет как палочковидную, так и разветвленную области, но не имеет центральной консервативной области и реплицируется в хлоропластах растительной клетки.
В отличие от вирусов, которые являются паразитами механизма трансляции хозяина, вироиды являются паразитами клеточных транскрипционных белков.
Бактериофаги

Существуют тысячи разновидностей фагов, каждый из которых может заразить только один тип или несколько близких типов бактерий или архей. Фаги классифицируются по ряду семейств вирусов; например:
Как и все вирусы, фаги являются простыми организмами, которые состоят из ядра генетического материала (нуклеиновой кислоты), окруженного капсидом белка. Нуклеиновая кислота может представлять собой либо ДНК, либо РНК, и может быть двухцепочечной или одноцепочечной.
Существует три основных структурных формы фага:
- Икосаэдрическая (20-сторонняя) головка с хвостом
- Икосаэдрическая головка без хвоста
- Нитевидная форма
Во время заражения фаг прикрепляется к бактерии и вставляет в нее свой генетический материал. После этого фаг обычно следует одному из двух жизненных циклов: литическому (вирулентному) или лизогенному (умеренному).
Литические, или вирулентные, фаги захватывают механизм клетки, чтобы скопировать компоненты фага. Затем они разрушают или лизируют клетку, высвобождая новые частицы фага.
Лизогенные, или умеренные, фаги включают свою нуклеиновую кислоту в хромосому клетки-хозяина и реплицируются с ней как единое целое, не разрушая клетку. При определенных условиях лизогенные фаги могут индуцироваться в соответствии с литическим циклом.
Существуют и другие жизненные циклы, в т.ч. псевдолизогенез и хроническая инфекция. При псевдолизогении бактериофаг проникает в клетку, но не использует механизм репликации клеток и не интегрируется в геном хозяина, просто как бы прячется внутри бактерии, не нанося ей никакого вреда. Псевдолизогенез возникает, когда клетка-хозяин сталкивается с неблагоприятными условиями роста и, по-видимому, играет важную роль в выживании фага, обеспечивая сохранение генома фага до тех пор, пока условия роста хозяина снова не станут благоприятными.
При хронической инфекции новые фаговые частицы образуются непрерывно и длительно, но без явного уничтожения клеток.
Вскоре после открытия фаги начали использовать для лечения бактериальных заболеваний человека, таких как бубонная чума и холера. Но фаговая терапия тогда не была успешной, и после открытия антибиотиков в 1940-х годах она была практически заброшена. Однако с появлением устойчивых к антибиотикам бактерий терапевтическому потенциалу фагов уделяется все больше внимания.
Наше время с антибиотиками заканчивается. В 2016 году женщина в штате Невада умерла от бактериальной инфекции, вызванной Klebsiella pneumoniae, которая была устойчивой ко всем известным антибиотикам. Бактерии, устойчивые к колистину, антибиотику последней инстанции, были обнаружены на свинофермах в Китае. В настоящее время бактерии приспосабливаются к антибиотикам быстрее, чем когда-либо.
Покажите ножницы которыми вирусы разрезают молекулу РНК что бы встроиться для мутации.Может что нибудь придумаете другое.К примеру деление цепочка аминокислот получив энергию из вне как одноименные заряды распадается на две. К каждой соединятся только те какие были ранее (другие проскочат мимо),казалось бы копии,но внутренняя энергия разная(уменьшается увеличивается) поэтому распад и создание. Вся химия углерода на этом построена 1000 орган соединений создает у других хим элементов этого свойства нет. Иммунная система делает накладку(интерференция)с помощью энергии интерферонов пытаясь разрушить цепочку РНК вируса.Надо помочь организму но не вакциной(вирус быстро мутирует)
Перед репликацией вирусная нуклеиновая кислота освобождается от капсида и оболочки. Например, у бактериофага Т4 капсид остается снаружи клетки-хозяина; у других капсид или оболочка отделяются от нуклеиновой кислоты внутри клетки; в третьем случае они могут растворяться ферментами клетки-хозяина. У некоторых вирусов часть белков должна внедриться в заражаемую клетку в комплексе с нуклеиновой кислотой, поскольку многие из них представляют собой полимеразы, которые необходимы для репликации вируса. Когда геном вируса освободится от капсида или оболочки внутри клетки-хозяина, дальнейшие события могут развиваться двумя путями.
2. Геном вируса встраивается в геном клетки-хозяина. Некоторые бактериофаги, например, имеют фрагменты ДНК, гомологичные участкам бактериального генома. Данные вирусы могут встраиваться в геном клеток путем рекомбинации, и в этом случае их называют умеренными фагами, поскольку лизиса клеток не происходит. Встроенный вирус называют профагом. Вирусы, участвующие в трансдукции, являются умеренными фагами. К умеренным вирусам относятся и некоторые вирусы животных, однако среди вирусов растений подобные случаи неизвестны.
У ДНК-содержащих вирусов, например, вируса осповакцины, вызывающего болезнь скота, ДНК направляет синтез мРНК, которая в свою очередь служит матрицей для синтеза различных белков. ДНК может быть одно- или двухцепочечной. У большинства РНК-содержащих вирусов, например, вируса табачной мозаики, геном представлен одноцепочечной молекулой. На ней в клетке-хозяине образуется комплементарная цепь, которая служит матрицей для синтеза новых РНК. Они связываются с рибосомами клетки и функционируют как информационная РНК, обеспечивая синтез ферментов и белков вирусной оболочки.
Одноцепочечные РНК-содержащие вирусы подразделяют на вирусы с позитивным (плюс-нитевые) и негативным (минус- нитевые) геномами. В первом случае РНК функционирует как матричная, во втором случае на ней образуется комплементарная цепь, которая служит матрицей для синтеза мРНК вируса. В свою очередь плюс-нитевые РНК-содержащие вирусы подразделяются на две группы в зависимости оттого, имеется ли у них оболочка. Например, вирус табачной мозаики не имеет оболочки. К безоболочечным РНК-содержащим вирусам с позитивным геномом относятся вирусы полиомиелита и ящура. Более того, к этому типу относится около одной трети вирусов, вызывающих респираторные заболевания человека.
К оболочечным РНК-содержащим вирусам с позитивным геномом относятся арбовирусы, переносимые членистоногими и вызывающие многие болезни, особенно в тропиках. Желтая лихорадка передается, например, от инфицированных обезьян к человеку с помощью москитов.
Минус-нитевые РНК-содержащие вирусы вызывают такие заболевания, как бешенство, корь, свинка, болезнь Ньюкасла домашних птиц и чума многих животных. Все перечисленные вирусы, кроме вируса бешенства, относятся к группе парамиксо вирусов и имеют сложное строение. Они покрыты оболочкой, имеют диаметр 300 нм и внутри аналогичны вирусу табачной мозаики, у которого вокруг РНК по спирали располагаются белковые субъединицы. Вирус гриппа относится к минус-нитевым РНК-содержащим вирусам; геном его представлен одноцепочечной РНК. К вирусам с двухцепочечной РНК относится вирус раневых опухолей растений (рис. 12-8), имеющий необычный процесс репликации и переносимый цикадками.
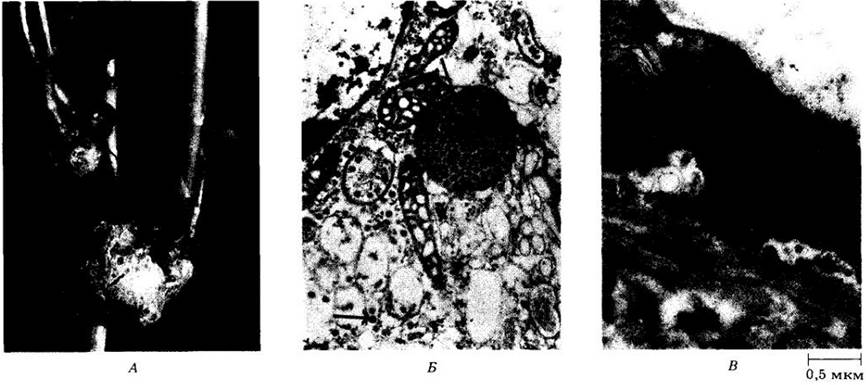
Рис. 12-8. А. Опухоли донника белого ( Melilotus alba), вызванные вирусом раневых опухолей. Б. Частицы вируса раневых опухолей (отменены стрелками) на электронной микрофотографии клетки растения-хозяина. В. Цикадка (Agallia constricta ) — переносчик вируса раневых опухолей. Электронная микрофотография эпидермальной клетки Agallia. Вверху слева видны многочисленные вирусные частицы, скопления которых напоминают соты. Внизу справа видны отдельные вирусные частицы. Вирус раневых опухолей, подобно многим вирусам растений, содержит двухцепочечную РНК
Две группы ДНК-содержащих вирусов — каулимовирусы и 1«минивирусы — поражают растения. Молекулярные свойства геминивирусов необычны; они представлены двумя вирионами, имеющими структуру несовершенных икосаэдров. В каждом вирионе заключена одноцепочечная молекула ДНК, причем для развития инфекции необходимы обе молекулы. Золотая мозаика бобов вызывается геминивирусами, которые переносятся белокрылками. Другая болезнь — полосатость кукурузы — вызывается вирусом, который распространяется цикадками.
Вирус мозаики цветной капусты переносится тлями; он относится к группе каулимовирусов, содержащих двухцепочечную ДНК. Частицы этих вирусов имеют икосаэдрическую форму. Разрывы в молекуле ДНК препятствуют образованию двойной спирали по всей ее длине. Каулимовирусы интенсивно изучаются, поскольку они могут быть переносчиками желаемых генов в растения.
Более тысячи известных заболеваний растений вызываются вирусами, относящимися примерно к 100 различным типам (рис. 12-8). Вирусные болезни растений, как правило, распространяются с помощью беспозвоночных — насекомых или нематод. Сосущие насекомые, такие, как тли и цикадки, переносят вирус вместе с соком, который извлекают из флоэмы или клеток эпидермиса. Некоторые вирусы размножаются в переносчике так же хорошо, как и в клетках инфицированного растения.
Некрозы характеризуются появлением участков мертвых тканей; в случае мозаичных болезней листья или другие части растения покрываются пятнами; оба эти широко распространенные заболевания вызываются вирусами. При мозаичной болезни на листьях или других зеленых частях растения появляются светло-зеленые и желтые маленькие крапинки или большие полосы. Иногда все инфицированное растение может быть более светлым, чем здоровое. Желтые пятна или окаймление листьев некоторых сельскохозяйственных культур вызываются вирусами, пестрая окраска цветов —тоже результат вирусной инфекции, передающейся от поколения к поколению (рис. 12-9).
Рис. 12-9. Пестрые цветки тюльпана. Подобная окраска вызвана вирусом

Вирусы мозаики преимущественно поражают ткани паренхимы, уменьшая или сводя к нулю количество хлоропластов. Другие накапливаются в богатом сахарами соке флоэмы и могут приводить к гибели ее клеток. Вирусные болезни в значительной степени снижают продуктивность зерновых культур во всем мире, причем они поражают огромное число видов. Безвирусные растения можно получать с помощью культуры ткани; при этом кончик меристематической ткани, не содержащий вируса, отделяют от родительского растения и культивируют in vitro , где он дает начало новому растению. Данный метод позволил увеличить урожай картофеля и ревеня.
Недавно сделано интересное открытие, касающееся вируса некротической мозаики риса, который при заражении растения подавляет его рост. С. К. Гхош из Индийского центрального института научных исследований в 1982 г. обнаружил, что другие растения (например, джут — источник грубых волокон для мешков и канатов) лучше растут, когда поражены вирусом, чем в здоровом состоянии. Этому явлению еще нет объяснения. Данный пример свидетельствует о сложной природе взаимодействия между вирусом и растением, но логично предположить, что в будущем подобные случаи будут выгодно использованы в экономическом отношении.
Вирусы растений, как правило, содержат РНК, за исключением каулимовирусов и геминивирусов (см. с. 179). В большинстве случаев капсид вирусов растений состоит из одного типа белка (комовирусов — из двух). Наиболее характерной особенностью PHК-содержащих вирусов растений можно считать часто фрагментированный геном. Препараты некоторых вирусов содержат несколько идентичных внешне типов вирусных частиц. Каждая частица имеет свой фрагмент РНК. Геном многих вирусов представлен двумя — четырьмя фрагментами РНК различного размера, каждый из которых заключен в отдельный капсид, так что вирусные частицы могут быть гетерогенными по форме и плотности. Пути взаимодействия фрагментов РНК, обеспечивающие инфекционность, неодинаковы у различных групп вирусов. У плюс-нитевых РНК-содержащих вирусов репликация происходит в два этапа: сначала на матрице исходной РНК образуется минус-нить; а затем на вновь образованной матрице синтезируется плюс-нить (мРНК).
Биологическая библиотека - материалы для студентов, учителей, учеников и их родителей.
Наш сайт не претендует на авторство размещенных материалов. Мы только конвертируем в удобный формат материалы, которые находятся в открытом доступе и присланные нашими посетителями.
Если вы являетесь обладателем авторского права на любой размещенный у нас материал и намерены удалить его или получить ссылки на место коммерческого размещения материалов, обратитесь для согласования к администратору сайта.
Разрешается копировать материалы с обязательной гипертекстовой ссылкой на сайт, будьте благодарными мы затратили много усилий чтобы привести информацию в удобный вид.
Под репликацией подразумевают процесс внедрения вирусов в восприимчивые к ним клетки, воспроизведение их генетического материала и белков, сборки и высвобождения способного к инфицированию потомства. Разнообразие вирусов, а именно их структуры и типа генетического материала, проявляется в целом ряде отличных друг от друга вариантов репликации. Первая стадия проникновения вирусов в клетки-мишени начинается с абсорбции вирусных частиц и заканчивается появлением вновь сформированного, способного к дальнейшему инфицированию поколения вирусов. Эта стадия, называемая часто эклипсным периодом (периодом затмения), продолжается от 1—5 ч у пикорнавирусов, тогавирусов, рабдовирусов, ортомиксовирусов, герпесвирусов и до 8—14 ч у аденовирусов, паповавирусов. В этот период происходит резкое снижение количества активных вирусов, поступающих из разрушенных клеток.
Адсорбция представляет собой, по всей видимости, первоначально обратимый процесс, являющийся результатом случайных столкновений вирусов с клетками-мишенями. Было установлено, что лишь в одном случае из 10 3 —10 4 таких столкновений происходит более тесное их связывание (склеивание). Связывание облегчается соответствующими ионными условиями и рН.
Однако этот процесс в основном не зависит от температуры и не требует затрат энергии. Адсорбция вируса на клетке-мишени подразумевает специфическое соединение вирусных белков с рецепторами клеточной мембраны (этот процесс также называется связыванием). У ряда вирусов были идентифицированы структуры, участвующие в процессе связывания вируса с клеткой. У вирусов, заключенных в конверты, белком связывания в типичных случаях служит один из пиков, находящихся на внешней поверхности вирусной оболочки, как, например, гемагглютинины (ГА) вирусов гриппа. Некоторые вирусы, имеющие оболочку, такие как герпесвирусы и вакцинии, содержат несколько типов белков, связывающихся с клетками. У вирусов без оболочки в качестве белков, связывающихся с клетками, часто выступают поверхностные полипептиды, такие как волокнистый белок аденовирусов и гемагглютинин (сигма 1) реовирусов.
Точная природа клеточных рецепторов животных вирусов установлена только для нескольких специфических видов. Даже если специфические рецепторы неизвестны, то с помощью исследований по конкурентному связыванию можно установить семейства или классы вирусных рецепторов. Вирусы одного вида, но разных серотипов могут конкурировать за рецепторы одного и того же класса (например, полиовирусы серотипов 1, 2, 3) или за рецепторы, принадлежащие к различным классам (например, риновирусы 2 и 14). Вирусы разных семейств (например, вирусы Коксаки В3 и аденовирусы 2) могут также конкурентно связываться с рецепторами одного и того же класса. Проведенные исследования по изучению процесса связывания показали, что на одной клетке имеется в целом 10 4 —10 6 мест для связывания с вирусом (рецепторы).
После того как присоединение вируса к клетке произошло, весь вирус или его субструктура, содержащая геном, и какая-либо вирусная полимераза, необходимая для первичной транскрипции, должны быть перенесены через плазматическую мембрану клетки. Скорость проникновения их различна и зависит от природы вируса, типа инфицируемой клетки и факторов окружающей среды, таких как температура. Некоторые не имеющие оболочки вирусы, такие как полиовирус и реовирус, вовлекаются в процесс эндоцитоза, регулируемого рецепторами (виропексис), в результате чего они попадают в цитоплазму внутри эндоцитотического пузырька. Другие не имеющие оболочки вирусы способны непосредственно проникать через плазматическую мембрану и находятся в цитоплазме в свободном состоянии без транспортных эндоцитотических пузырьков.
Проникновение в клетку вирусов, покрытых оболочкой, также может осуществляться двумя путями. Примером первого служит вирус леса Семлики (ВЛС). ВЛС, являющийся представителем семейства тогавирусов, связывается со специфическими рецепторами клеточной поверхности, которые затем группируются в определенных местах плазматической мембраны (прикрытые колодцы), а затем поступают внутрь клетки посредством рецепторного эндоцитоза. Впоследствии они появляются в клеточной цитоплазме ,в пузырьках, покрытых клатрином. Слияние оболочки вируса с эндосомальной мембраной вызывает высвобождение вирусного капсида в цитоплазму. Второй механизм проникновения покрытых вирусов имеет место у парамиксовирусов (например, Сендай). Оболочка вируса сливается непосредственно с клеточной плазматической мембраной, и вирусный нуклеокапсид в свободном виде поступает в цитоплазму.
Раздевание представляет собой процесс удаления или распада части или всего вирусного белкового капсида на этапе подготовки вирусного генома к транскрипции и трансляции. Во многих случаях проникновение и раздевание являются частью одного процесса. У некоторых пикорнавирусов, например, могут происходить изменения структуры и функции капсида, нарушаться его целостность и исчезать внутренние белки по мере продвижения вируса через плазматическую мембрану. Структурные изменения, сопровождаемые утратой белка, облегчают внедрение вирусной РНК в цитоплазму.
Вирусы, не покрытые оболочкой, например аденовирусы, поступая в эндосомы, индуцируют слияние лизосом с эндосомой, в результате чего их капсид удаляется под воздействием лизосомальных ферментов. В случае реовирусов интраэндосомальные протеазы последовательно убирают три внешних белка капсида, в результате чего образуется субвирусная частица. Этот процесс ведет к активации вирусной транскриптазы. Раздевание поксвирусов, таких как вирус осповакцины, на первых этапах заключается в разрушении внешнего белкового покрытия с помощью интраэндосомальных ферментов, а затем и белков оставшегося ядра. В конечном итоге высвобождается вирусная ДНК. Данный этап, видимо, требует синтеза вирусспецифического раздевающего белка.
Транскрипция вирусного генома в иРНК и последующая трансляция иРНК в белок могут проходить по целому ряду путей. В целом для эукариотических клеток необходимо, чтобы иРНК имели один-единственный начальный участок для трансляции белка (т. е. они моноцистроничны). Путь у вирусов, содержащих иРНК, заключается в трансляции ее в большой первичный полипротеин, который затем расщепляется с образованием нескольких вирусных протеинов. Примером такого пути могут быть вирусы, нуклеиновая кислота которых находится в форме (+)-полярной, односпиральной РНК (осРНК) и выступает в роли иРНК, в частности пикорнавирусы и тогавирусы. Она связывается с большими полирибосомами и полностью <53)транслируется, образуя один большой полипротеин, который затем расщепляется, проходя несколько стадий, что сопровождается появлением неструктурных, ядерных и капсидных белков.
У тогавирусов вирусная РНК выступает в роли и иРНК, формируя полипротеин, расщепляющийся с последующим формированием неструктурных протеинов, необходимых для репликации РНК. Затем вирусная РНК транскрибируется в (—) РНК, служащую матрицей длины генома, с которой копируются два основных типа (+) РНК. Между альфа-вирусами и флавовирусами семейства тогавирусов существуют большие различия. Например, длина иРНК флавовирусов соответствует длине генома, в то время как размеры иРНК альфа-вирусов меньше генома. Кроме того, гены структурных белков флавовирусов локализуются на 5^конце вирусного генома, в то время как гены структурных белков альфа-вирусов находятся на его 3^конце.
Как у пикорнавирусов, так и у тогавирусов вирусная закодированная РНК-полимераза синтезирует комплементарную РНК, используя в качестве матрицы РНК генома. Вновь синтезированная РНК в свою очередь служит матрицей для синтеза последующих РНК генома. Новые геномные РНК могут выступать в роли иРНК или предшественников РНК для следующих поколений вирусов.
Вирусы, содержащие линейную или сегментарную РНК, продуцируют не одну большую молекулу иРНК, а специфические иРНК для каждого вирусного белка. Для создания иРНК из геномной РНК необходим фермент транскриптаза, содержащийся в вирионе (вирусная полимераза). Наличие множества иРНК позволяет регулировать количество каждого синтезируемого белка. Один участок геномной РНК может иметь множество считывающих рамок, каждая из которых транскрибируется в уникальную иРНК. А она в свою очередь транслируется в отдельный белок. Геномная (—) осРНК реплицируется с участием (+) осРНК-посредника, которая затем служит матрицей для синтеза большого количества (—) осРНК генома.
Реовирусы содержат РНК-зависимую РНК-полимеразу, транскрибирующую (+) осРНК из (—)-трабекул каждого сегмента двуспиральной (дс) РНК. Эти (+)осРНК с силой выбрасываются из ядра вируса через каналы в ядерных пиках и служат в качестве моноцистрональных иРНК для трансляции в вирусные белки. Вирусная РНК-полимераза также синтезирует (+)осРНК, которые в свою очередь служат матрицами для комплементарных (—)-трабекул во время репликации вирусного генома.
У ретровирусов единый путь репликации. Вирусная (+) осРНК служит матрицей для вирусной РНК-зависимой ДНК-полимеразы (обратная транскриптаза) и первичных транспортных РНК (тРНК). Получается копия осДНК, которая первоначально соединена водородной связью с ее комплементарной (+) осРНК. Вирусная закодированная рибонуклеаза расшифровывает осРНК, после чего синтезируется комплементарная цепь ДНК. Затем дсДНК интегрируется в хромосомную ДНК ядра клетки-хозяина. Транскрипция этой интегрированной вирусной ДНК находится под контролем транскриптаз клетки-хозяина.
ДНК-содержащие вирусы способны использовать пути, сходные с теми, которые описаны для эукариотических клеток при их репликации во время литической инфекции. Паповавирусам, аденовирусам и герпесвирусам свойствен такой путь репликации, при котором транскрипция вирусной ДНК в иРНК происходит в ядре клетки-хозяина и зависит от ферментов последней. У паповавирусов (т. е. SV40) первичными протеинами, образующимися после внедрения, являются Т-антигены (опухолевые антигены, или ранние белки). Некоторые из Т-антигенных белков способны взаимодействовать с дсДНК вирусного генома, присоединяясь вблизи от мест, запускающих репликацию ДНК, что облегчает репликацию последней. Впоследствии транскрибируются иРНК, в которых закодированы полипептиды капсида (поздние белки). Все ранние иРНК являются производными какой-либо одной из двух цепей вирусной ДНК (называемой Р, или ранней, цепью), а поздние иРНК — другой (П, или поздней, цепью). Аденовирусы также имеют ранние и поздние гены, но они располагаются не на разных цепях вирусной ДНК, а беспорядочно на обеих.
При репликации как паповавирусов, так и аденовирусов ранние белки играют главным образом регулирующую роль и часто проявляют плейотропное действие. Поздние белки включают структурные протеины. Отдельные иРНК как для ранних, так и для поздних белков часто комплементарны сегментам, находящимся на разных участках вирусной ДНК. Это указывает на значительное сращивание участков вирусной ДНК с удалением участков, находящихся между комплементарными сегментами, в последующем. Во многих случаях иРНК синтезируются на основе накладывающихся друг на друга участков вирусной ДНК. Такой тип наслаивания позволяет уменьшить количество вирусной ДНК, необходимой для кодирования вирусных белков.
Репликация вирусной ДНК паповавирусов и аденовирусов несколько различается. В обоих случаях необходимо участие ДНК-полимеразы, выделенной из клеток хозяина. Репликация ДНК у паповавирусов начинается в каком-то одном месте и затем распространяется в двух направлениях вдоль циркулярной дсДНК до тех пор, пока обе репликационные вилки не встретятся. Синтез ДНК протекает не бесконечно. Небольшие вновь синтезированные фрагменты ДНК позже соединяются вместе по меньшей мере на одной из двух цепей ДНК. Репликация дсДНК у аденовирусов облегчается ее линейной, а не циркулярной организацией. Репликация каждой отдельной комплементарной цепи ДНК происходит независимо, а вновь синтезированные цепи могут инициировать следующий круг репликации.
У герпесвирусов вирусная ДНК первоначально транспортируется в клеточное ядро, где происходит транскрипция и репликация. Самые ранние появляющиеся протеины выполняют регуляторную функцию. Протеины, появившиеся позднее, участвуют в синтезе вирусной ДНК. Белки, сформированные позже всех, имеют структурную природу.
Поксвирусы являются наиболее сложными из всех известных вирусов животных. Соответственно их репликационный цикл также сложен. Все первичные стадии транскрипции и трансляции протекают, по всей видимости, в цитоплазме клетки-хозяина. Для этого требуется, чтобы вирус имел собственную ДНК-зависимую РНК-полимеразу, инициирующую транскрипцию. Один из вирус -закодированных ранних белков отвечает за вторую стадию раздевания, после чего вирусная ДНК становится полностью доступной для транскрипции и репликации. Репликация, транскрипция, а позже и сборка вирусов — все это происходит на фабриках, находящихся в цитоплазме клетки-хозяина и активируемых вирусом. Остаточные группы вирусспецифических белков можно обнаружить в инфицированных клетках. К ранним белкам относятся некоторые ферменты (например, ДНК-полимераза и тимидинкиназа) и структурные белки. По мере прогрессирования инфекционного процесса начинается репликация ДНК, синтез ранних неструктурных белков прекращается и начинается синтез поздних белков, многие из которых являются структурными, другие — ферментами и протеинами, участвующими в сборке вирусов.
После окончания репликации вирусного генома и синтеза вирусных белков должны начаться сборка интактных вирионов и их выделение из клетки-хозяина. Сборка вирусов, не имеющих оболочки и нуклеокапсидов вирусных оболочек часто протекает по типу кристаллизации, который зависит от самосборки вирусных капсомеров.
В большинстве случаев вирусы, не имеющие оболочки, накапливаются внутри пораженной клетки и выделяются во внешнюю среду после гибели клетки. К разрушению клетки приводит следующая цепь событий: угнетение синтеза белков клетки-хозяина, ее липидов и нуклеиновых кислот; дезорганизация цитоскелета клетки-хозяина; изменение структуры мембраны клетки-хозяина. Разрыв мембран приводит к повышению клеточной проницаемости и высвобождению протеолитических ферментов из лизосом. Невозможность пополнить запасы высокоэнергетических молекулярных структур подавляет функцию насосов, обеспечивающих движение ионных потоков, и нарушает выведение продуктов распада и поступление необходимых питательных веществ.
Вирусы, имеющие оболочку, высвобождаются из инфицированной клетки, как правило, почкованием. Этот процесс может окончиться гибелью клетки. Во всех случаях вирусспецифические белки встраиваются в мембраны клеток хозяина, вызывая их структурную перестройку, смещая некоторые нормальные белковые компоненты. В последующем вирусные капсиды могут связываться с вирусспецифическими белками матрикса, выстилающими мембрану со стороны цитоплазмы в поврежденных участках. Капсиды вирусов, имеющих самые маленькие размеры и покрытые оболочкой, связываются с интрацитоплазматическими областями вирусных белков, встроенных в мембрану клетки-хозяина, а не с матриксными белками (тогавирусы).
Читайте также:
