
Репликации днк-содержащих ретроидных вирусов
Сайт СТУДОПЕДИЯ проводит ОПРОС! Прими участие :) - нам важно ваше мнение.
Варианта вирусных частиц
РЕТРОИДНЫЕ (ПАРАРЕТРО) ВИРУСЫ.
( СОДЕРЖАТ ДНК, НО КОДИРУЮТ ОБРАТНУЮ ТРАНСКРИПТАЗУ)
Эти вирусы используют матричную РНК, построенную на матрице ДНК не только для синтеза белков, но и для репликации дочерних геномов вируса.
Общие свойства параретровирусов:
Кольцевая частично двунитевая геномная ДНК: т е есть одно кольцо полностью и второе, комплементарное ему, незамкнутое и не соответствующее полностью по длине (-)ДНК.
• Небольшой размер генома – примерно 3000-8000 нуклеотидов.
• Обратная транскриптаза (RT) присутствует в составе вирионов.
• Нет необходимости встраиваться в хозяйскую хромосому для успешной репликации. Репликация происходит в цитоплазме, ДНК при этом образует в ядре зараженной клетки эписому – сверхспирализованную замкнутую 2-цепочечную структуру, которая необходима для транскрипции.
• Группа VII ( по Балтимору)
Обнаружены и у животных и у растений. Организованы в 2 семейства:
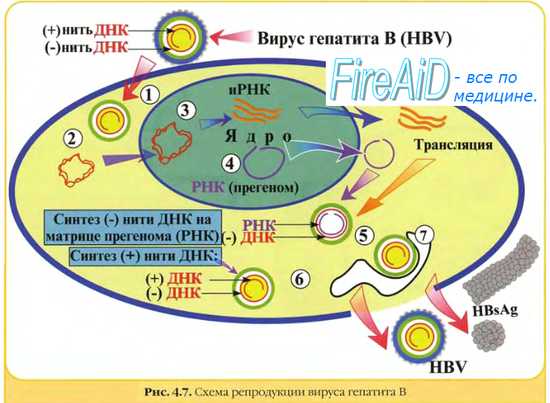
• Семейство Hepadnaviridae,(заражают только гепатоциты), передаются через кровь, половым путем и вертикально (от матери к ребенку)
род Orthohepadnavirus (млекопитающие) и
род Avihepadnavirus (птицы).
HBV, вирус гепатита B. Вирионы – икосаэдр,
покрытый липидной оболочкой.
• Семейство Caulimoviridae, 7 родов. CaMV,
cauliflower mosaic virus, вирус мозаики цветной капусты. Икосаэдрической или бацилловидной формы, липидная оболочка
отсутствует, заражают растения
род Badnavirus
• 1 – инфекционные частицы Дейна
42 нм;(70% приходится на белок S). Австралийский ген, или HВsAgв крови говорит о носительстве вируса гепатита В и о риске образования карциномы печени.
К 5 концу (-)цепи присоединен белок Р (полимераза, по сути ревертаза). На 5 конце (+) цепи находится фрагмент РНК, т о + и – цепи используют различные затравки для своей репликации.
• 2 – неинфекционные сферические 20нм, это HВsAg (почти только S белок) и мембрана и все. Они накапливаютя, 10 тыс – 1 млн на 1 частицу Дейна, полезно для диагностирования.
• 3 -нитевидные различной длины диаметром 20нм.
Вирионы 2 и 3 содержат только поверхностный антиген HBsAg (S-белок) без геномной ДНК и белка сердцевины (core) и
присутствуют в большом избытке.
Еще раз о белках: HВsAg
3 поверхностных антигена – L(39кДа),M(31кДа),S(24кДа).
• Антиген S – основной компонент оболочки: L и M составляют 2% и 5-10% от общего количества.
• Антиген L необходим для связывания с клеточным рецептором и для созревания вирусных частиц. Взаимодействует с коровой частью вириона и приводит к его переносу через мембрану эндоплазматической сети.
· После попадания в ядро неполная (+) нить ДНК достраивается до полного размера, рибонуклеотидный праймер удаляется с ее 5 конца.
· С 5 конца (-) ДНК удаляется молекула полимеразы
· Все разрывы в молекулах лигируются с помощью ферментов клетки
· Двуцепочечная ДНК вируса гепатита В сверхспирализуется тоже с помощью ферментов клетки, находящихся в ядре
· Итак, в виде такой эписомы ДНК служит матрицей для транскрипции
· Эта эписома в ядре может реплицироваться, но в незначительном количестве, основная репликация вирусного генома будет происходить в цитоплазме
ORF – открытая рамка считывания, HВsAg – зеленый
Есть несколько ORF:
ORF P - revertase.
ORF S/preS - гликопротеиды оболочки
ORF C - белок нуклеокабпсида, С-белок
ORF X HBX, 17 кда, неструктурный белок, трансактиватор экспрессии
генов, основной фактор онкогенного потенциала гепатита Б. Геном
вируса гепатита Б 3.5 кб - небольшой. Здесь опять используется
перекрывание генов - все гены перекрываются. Транскрипция на
эписоме идет с помощью клеточной РНК полимеразы. ранскрибируется
4 мРНК, все кэпировано, полиаденилировано.Промоторы, с которых идет образование этих матричных РНК ,находятся в разных участках генома, а 3 концы идентичны. Начинается их синтез в разных точках, но оканчивается в одной, они котерминальны.
С самой маленькой м-РНК транслируется белок Х.
АУГ белка S находится ближе всего к 5 концу и кэпу, а АУГ М белка находится чуть дальше в другой рамке считывания, а т к эукариотические рибосомы предпочитают 1ый АУГ , белка S на капливается очень много, а белок М инициируется от случая к случаю и его образуется меньше. Аналогично происходит при трансляции прегеномной РНК.
Итак, вот эта РНК является матрицей для белка кора и ревертазы и матрицей для репликации белка.
Оказывается, что длина прегеномной РНК больше самого генома! С нее транслируется тоже 2 белка НВс и полимераза.
В вирусную частицу входит около 200 копий корового белка и всего одна молекула полимеразы. АУГ С белка находится ближе к 5 концу и происходит нормальная кэп-зависимая инициация, эффективная, а АУГ полимеразы находится на некотором расстоянии, тоже со сбоем рамки, и полимеразы образуется очень мало, где-то 1 молекула на вирусную частицу.
Для того ,чтобы начать процесс обратной транскрипции обратная транскриптаза взаимодействует с прегеномной РНК и блокирует трансляцию прегеномной РНК. Трансляции не происходит и начинается формирование кора, в котором и происходит обратная транскрипция. пгРНК длиннее, т к участок геномной РНК при ее транскрипции прочитывается дважды, что приводит к образованию повторов на 5 и 3 концах пг РНК. Повторы довольно крупные, примерно 200нуклеотидов(R-повторы) внутри них более короткие повторы - DR1 (всего несколько нуклеотидов). Кроме того, на 5' 3'-концевого R повтора есть DR2 повтор. Рядом с короткими повторами DR1 есть эпсилон-шпилька на 3 конце. Первый акт при начале репликации – связывание полимеразы со структурой эпсилон инициирует инкапсидирование РНК в капсидный белок и формирование сердцевины вириона и одновременно инициируется процесс обратной транскрипции с образованием короткого фрагмента (-)цепи ДНК (всего несколько нуклеотидов), при этом молекула полимеразы выполняет роль затравки, через тирозин ковалентно связывается с первыми нуклеотидами ДНК, СВЯЗЬ ОСТАЕТСЯ НА ВСЕ ВРЕМЯ И ИМЕННО В ТАКОМ ВИДЕ ПОЛИМЕРАЗА ПОПАДАЕТ В ВИРУСНУЮ ЧАСТИЦУ.
| Р.прямоугольники с циферками - это повторы, по порядку слева направо: 1, 2, 1 |
Итак, на пгРНК строится участок, комплементарный боксу DR1, происходит прыжок полимеразы ,
она транслоцируется в область другого DR1
повтора. Обратная транскриптаза обладает РНКаза Н активностью и постепенно удаляет РНК в гетеродуплексе РНК-ДНК. По непонятным причинам остается небольшой участок прегеномной РНК, ее 5 конец.
Происходит второй прыжок полимеразы между DR2 и DR1, (они комплементарны). Т о образуется свободный 3 конец, необходимый для синтеза (+)ДНК, т е затравкой для синтеза (+)цепи ДНК является фрагмент прегеномной РНК. Ну и происходит синтез этого небольшого участка (+)ДНК с помощью полимеразы. Дальше за счет той же комплементарности (-)ДНК образует кольцо, благодаря этому полимераза переходит на 3 конец (-)цепи и продолжается синтез (+)ДНК. (+)цепь наращивается полимеразой до тех пор, пока сердцевина вириона, в которой идет этот процесс, не связывается с L белком на мембране (он находится в составе мембранной эндоплазматической сети), это связывание ведет к небедленной остановке синтеза (+) нити. Процесс может быть растянут по времени, поэтому разные вирионы могут иметь (+)ДНК разной длины, но никогда (+)ДНК не успевает сделать полный круг. А кусочек пгРНК остается ковалентно связанным с (+)ДНК вируса и в составе вириона.
Итак, репликация идет в цитоплазме, внутри коровой частицы,
которая параллельно формируется из продуктов трансляции. В образовании этих коровых частиц участвуют шапероны клетки, более того часто сами входят в состав созревших вирусных частиц (БТШ 90).
Суммируем: частица взаимодействует с неизвестным пока рецептором, ДНК идет в ядро, там происходит изменение 5' концов, ДНК достраивается, лигируется и образуется эписома, с нее идет транскрипция, затем с прегеномной РНК в цитоплазме связывается обратная транскриптаза и идет
репликация, одноврменно при участии клеточных шаперонов
образуется кор, он связывается с HBS L на ЭПР и репликация
преркащается, при этом +цепь не успевает достроиться. Важно, что
гепатит б распространен неравномерно, очень много его в Китае.
Заражается только один тип клеток, в патогенезе важна иммунная система, 95 процентов излечивается самостоятельно с помощью цитотоксических Т-лимфоцитов. При этом больные теряют часть печени - отсюда симптомы. У 5 процентов слабый иммунный ответ и они становятся хрониками. Сейчас в мире 300млн носителей. Это огромный резервуар и среди них 10 процентов
получают рак печени.
Почему вирус онкогенный? Геном вируса способен интегрировать в клеточный геном. И при этой случайной интеграции вирус нарушает транскрипцию клеточных генов, что очень часто приводит к потере контроля над клеточным делением. Есть другая версия: при интеграции ген Х никогда не теряется, он является транскрипционным активатором, помимо своих, активирует факторы транскрипции самой клетки, ну и, соответственно, белков, которые регулируют клеточные деления.
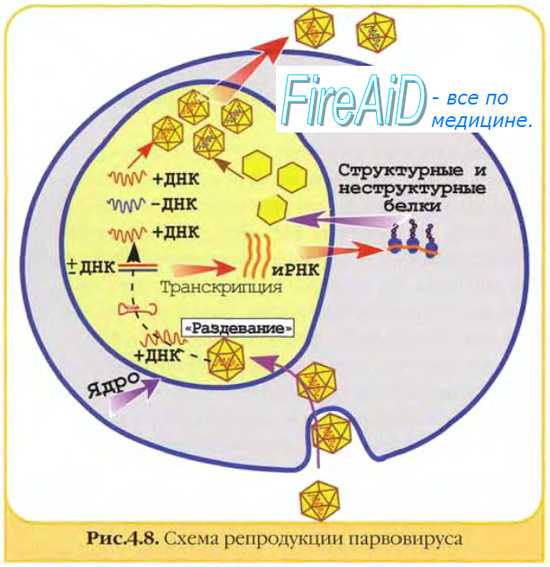
Семейство Parvoviridae
Около 50 представителей, образуют 2
подсемейства: Parvovirinae (позвоночные) и Densovirinae (насекомые).
• Геном – линейная ssДНК
• Группа II (по Балтимору)
• Различают автономные и дефектные или
ассоциированные парвовирусы (род Dependovirus, аденосателлит AAV2).
Наиболее известный парвовирус, заражающий человека – это парвовирус В-19, он вызывает инфекционную эритему, а у младенцев – пятую болезнь, размножается в крови, костном мозге, в тканях эмбриона, около 70% людей – носители этого вируса.
• Автономные парвовирусы размножаются только в S- фазе клеточного цикла, но не способны стимулировать деление зараженной клетки, они имеют все компоненты для самостоятельного размножения.
• Ассоциированным (дефектными) парвовирусам необходим помощник – адено- или герпесвирус, в его отсутствие способны встраиваться в геном
клетки-хозяина (провирус) и таким образом спасаются(очень четко, в 19 хромосому человека и поэтому к образованию опухолей не приводит). Помощник стимулирует транскрипцию клетки.
Дальше все обсуждаем на примере AAV2.
2 открыты рамки считывания (регулят и структур белки).
Проникают в различные типы клеток посредством эндоцитоза.. В кислой среде эндосомы вирионы претерпевают конформационные изменения,выходят в цитоплазму. Терминальный участок белка VP1 обладает активностью фосфолипазы и обеспечивает транспорт фирусных частиц через эндолизосому.
Оказавшись в цитоплазме, вирусные частицы транспортируются по микротрубочкам к ядру и проникают через ядерные поры (размер позволяет). В ядре ДНК освобождается от капсида и начинается репликация, она осуществляется ДНК-полимеразой клетки.
Прописная и заглавная буквы на картинке – это участки, комплементарные между собой. Благодаря вторичной структуре мол-лы не нужно никакой затравки, полимераза сразу же может начинать свою работу.
-Половина дочерних нитей ДНК аденосателлита имеет положительную, а другая – отрицательную полярность.
• Часть материнской молекулы входит в состав дочерней молекулы ДНК.
• Нуклеотидная последовательность концевых повторов у дочерних молекул ДНК отличается от последовательности родительской ДНК.
• Происходит инверсия нуклеотидной последовательности внутри концевого повтора.
• Структура концевой последовательности (инвертированный повтор) сохраняется
• Изменения не затрагивают смысловых нуклеотидных последовательностей (как кодирующих, так и регуляторных) и не влияют на репликацию вируса.
Вирусы, лишённые суперкапсида (например, аденовирусы) проникают в клетки путём виропексиса, а имеющие таковой (покс- и герпесвирусы) — за счёт слияния суперкапсида с клеточной мембраной. Репродуктивный цикл ДНК-содержащих вирусов включает раннюю и позднюю стадии (рис. 5-4). У крупных ДНК-вирусов имеется явное несоответствие между кодирующе ёмкостью генома и молекулярной массой вирусиндуцированных белков и белков, входящих состав вирионов. Например, у герпесвирусов лишь 15% ДНК кодирует все белки вирионов и их предшественников. Возможно, значительная часть генома содержит гены, кодирующие синтез ферментов и регуляторных белков. Папова-, адено- и герпесвирусы репродуцируются относительно однотипно, в то время как репродукция поксвирусов имеет некоторые особенности.
Регуляторные белки выполняют различные функции. При заражении клетки они блокируют синтез клеточных РНК, ДНК и белка и одновременно способствуют экспрессии вирусного генома, изменяя специфичность реагирования клеточных полимераз и полирибосом. Они так же запускают репликацию клеточной ДНК, модифицированной встроенными геномами ДНК содержащих вирусов и ретровирусов, то есть репликацию вирусных геномов. Вирусспецифические полимеразы. В репликацию вирусных геномов также вовлечены вирусоспецифические ДНК-полимеразы, участвующие в образовании молекул ДНК дочерних популяций.
Матричные белкинеобходимы для репликации нуклеиновых кислот и сборки дочерних популяций. Они образуют электронно-плотные скопления в клетке, известные как тельца включений (например, тельца Гварнери при натуральной оспе).
Паповавирусы и аденовирусы. Репродукция паповавирусов. Репродукция аденовирусов.
Адсорбция, проникновение и депротеинизация аналогичны таковым у РНК-содержащих вирусов, но у папова- и аденовирусов депротеинизация протекает в ядре, а у РНК-вирусов — в цитоплазме.
начальный период включает стадии адсорбции вируса на клетке, проникновения в клетку, дезинтеграции (депротеинизации) или "раздевания" вируса. Вирусная нуклеиновая кислота была доставлена в соответствующие клеточные структуры и под действием лизосомальных ферментов клетки освобождается от защитных белковых оболочек. В итоге формируется уникальная биологическая структура: инфицированная клетка содержит 2 генома (собственный и вирусный) и 1 синтетический аппарат (клеточный);
• после этого начинается вторая группа процессов репродукции вируса, включающая средний и заключительный периоды, во время которых происходят репрессия клеточного и экспрессия вирусного генома. Репрессию клеточного генома обеспечивают низкомолекулярные регуляторные белки типа гистонов, синтезируемые в любой клетке. При вирусной инфекции этот процесс усиливается, теперь клетка представляет собой структуру, в которой генетический аппарат представлен вирусным геномом, а синтетический аппарат — синтетическими системами клетки.
2. Дальнейшее течение событий в клетке направлено на репликацию вирусной нуклеиновой кислоты(синтез генетического материала для новых вирионов) и реализацию содержащейся в ней генетической информации (синтез белковых компонентов для новых вирионов). У ДНК-содержащих вирусов, как в прокариотиче-ских, так и в эукариотических клетках, репликация вирусной ДНК происходит при участии клеточной ДНК-зависимой ДНК-полимеразы. При этом у однонитевых ДНК-содержащих вирусов сначала образуется комплементарная нить — так называемая репликативная форма, которая служит матрицей для дочерних молекул ДНК.
3. Реализация генетической информации вируса, содержащейся в ДНК, происходит следующим образом: при участии ДНК-зависимой РНК-полимеразы синтезируются и-РНК, которые поступают на рибосомы клетки, где и синтезируются вирусспе-цифические белки. У двунитевых ДНК-содержащих вирусов, геном которых транскрибируется в цитоплазме клетки хозяина, это собственный геномный белок. Вирусы, геномы которых транскрибируются в ядре клетки, используют содержащуюся там клеточную ДНК-зависимую РНК-полимеразу.
У РНК-содержащих вирусов процессы репликацииих генома, транскрипции и трансляции генетической информации осуществляются иными путями. Репликация вирусных РНК, как минус-, так и плюс-нитей, осуществляется через репликативную форму РНК (комплементарную исходной), синтез которой обеспечивает РНК-зависимая РНК-полимераза — это геномный белок, который есть у всех РНК-содержащих вирусов. Репликативная форма РНК минус-нитевых вирусов (плюс-нить) служит не только матрицей для синтеза дочерних молекул вирусной РНК (минус-нитей), но и выполняет функции и-РНК, т. е. идет на рибосомы и обеспечивает синтез вирусных белков (трансляцию).
У плюс-нитевыхРНК-содержащих вирусов функцию трансляции выполняют ее копии, синтез которых осуществляется через репликативную форму (минус-нить) при участии вирусных РНК-зависимых РНК-полимераз.
У некоторых РНК-содержащих вирусов (реовирусы) имеется совершенно уникальный механизм транскрипции. Он обеспечивается специфическим вирусным ферментом — ревертазой (обратной транскриптазой) и называется обратной транскрипцией. Суть ее состоит в том, что вначале на матрице вирусной РНК при участии обратной транскрипции образуется транскрипт, представляющий собой одну нить ДНК. На нем с помощью клеточной ДНК-зависимой ДНК-полимеразы синтезируется ,вторая нить и формируется двунитевой ДНК-транскрипт. С него обычным путем через образование и-РНК происходит реализация информации вирусного генома.
Результатом описанных процессов репликации, транскрипции и трансляции является образование дочерних молекул вирусной нуклеиновой кислоты и вирусных белков, закодированных в геноме вируса.
После этого наступает третий, заключительный период взаимодействия вируса и клетки. Из структурных компонентов (нуклеиновых кислот и белков) на мембранах цитоплазматического ретикулума клетки собираются новые вирионы. Клетка, геном которой был репрессирован (подавлен), обычно гибнет. Вновь сформировавшиеся вирионы пассивно (в результате гибели клетки) или активно(путем почкования) покидают клетку и оказываются в окружающей ее среде.
Таким образом, синтез вирусных нуклеиновых кислот и белков и сборка новых вирионов происходят в определенной последовательности (разобщены во времени) и в разных структурах клетки (разобщен в пространстве), в связи с чем способ репродукции вирусов и был назван дизъюнктивным (разобщенным). При абортивной вирусной инфекции процесс взаимодействия вируса с клеткой по тем или иным причинам прерывается до того, как произошло подавление клеточного генома. Очевидно, что в этом случае генетическая информация вируса реализована не будет и репродукции вируса не происходит, а клетка сохраняет свои функции неизменными.
При латентной вирусной инфекции в клетке одновременно функционируют оба генома, а при вирус-индуцированных трансформациях вирусный геном становится частью клеточного, функционирует и наследуется вместе с ним.
Дата добавления: 2018-05-09 ; просмотров: 466 ;

Учебник предназначен для студентов, аспирантов и преподавателей высших медицинских учебных заведений, университетов, микробиологов всех специальностей и практических врачей.
5-е издание, исправленное и дополненное
Книга: Медицинская микробиология, иммунология и вирусология
Репликация вирусных геномов
Репликация вирусных геномов
Тип геномной вирусной ДНК определяет особенности ее репликации.
1. Двунитевая ДНК – репликация происходит по обычному механизму полуконсервативной репликации: нити разделяются, и на каждой из них достраивается комплементарная ей нить.
3. У вирусов, геном которых представлен однонитевой РНК, ее репликация происходит по следующей схеме: вначале на вирионной РНК (вРНК) синтезируются комплементарные ей РНК (кРНК). Этот процесс катализируется специфической РНК-репликазой I. Затем на кРНК синтезируется комплементарная ей, но идентичная исходной вирионная РНК (вРНК), этот процесс также катализируется специфической репликазой II. Таким образом, репликация идет по схеме (рис. 80. II):
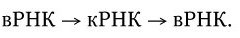
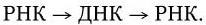
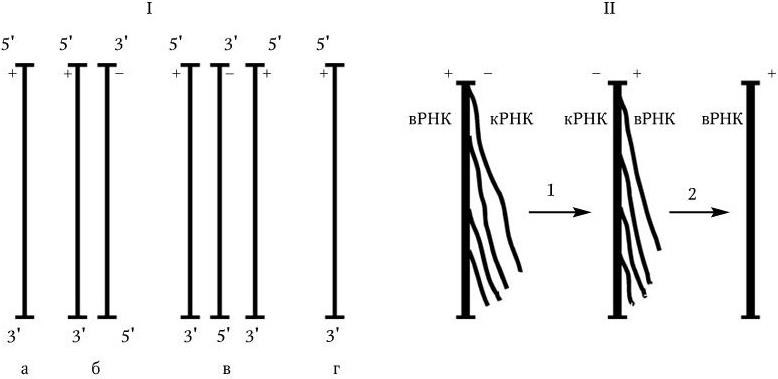
Рис. 80. Механизм репликации однонитевого вирусного ДНК-генома (I) и однонитевого вирусного РНК-генома (II):
а – геномная однонитевая ДНК; б – репликативная форма; в – промежуточная репликативная форма; г – однонитевая дочерняя геномная ДНК; вРНК – вирионная (геномная) РНК; кРНК – комплементарная РНК. Объяснение в тексте
5. Размножение вируса гепатита В также протекает с участием обратной транскриптазы, но вначале клеточная РНК-полимераза синтезирует на вирусной ДНК прегеномную РНК, после чего вирусная ревертаза синтезирует на ней минус-цепь ДНК, которая достраивается плюс-цепью ДНК. В виде двунитевой ДНК вирус интегрируется в хромосому клетки-хозяина, где на ней транскрибируется вирусная РНК.
Существуют некоторые общие закономерности размножения вирусов. Во-первых, все РНК-содержащие вирусы, кроме вирусов гриппа и ретровирусов, размножаются в цитоплазме. Для своего размножения вирусы гриппа А и В и ретровирусы проникают в ядро, что связано с особенностями поведения их генома. Во-вторых, размножение всех ДНК-содержащих вирусов, кроме вирусов оспы, протекает в ядре, где происходит транскрипция и репликация их геномных нуклеиновых кислот, и в цитоплазме, где происходит трансляция вирусных белков, их процессинг и морфогенез вирионов. Лишь размножение вирусов группы оспы происходит в цитоплазме клетки, поскольку они обладают собственными системами транскрипции.
Другая особенность размножения вирусов заключается в том, что их нуклеокапсидные белки синтезируются на свободных полирибосомах (не связанных с мембраной), а суперкапсидные белки – на рибосомах, ассоциированных с мембранами (на шероховатых мембранах). Кроме того, белки некоторых вирусов подвергаются протеолитическому процессингу и гликозилированию. Различают два типа протеолитического процессинга: каскадный и точечный.
При точечном протеолизе разрезанию подвергается один (реже несколько) из вирусных полипептидов. Разрезание происходит, как правило, в определенном участке полипептида. Такой тип протеолиза необходим для того, чтобы определенный белок вируса приобрел свою специфическую активность. Например, суперкапсидный белок вируса гриппа – гемагглютинин – разрезается на две субъединицы: боEльшую и меньшую. В результате меньшая субъединица приобретает способность сливаться с мембранами клетки-мишени и ее лизосомами. Благодаря этому вирус гриппа приобретает способность проникать в клетку. Такой точечный протеолиз наблюдается у ортомиксовирусов, парамиксовирусов, ротавирусов, вирусов группы оспы и др. Точечный протеолиз, как и каскадный, жизненно важен для вируса.
Наконец, еще одна особенность вирусов, обладающих суперкапсидом, заключается в том, что суперкапсидные белки подвергаются в ходе своей транспортировки на наружную поверхность клеточной мембраны гликозилированию.
Геном вирусов может быть представлен одноцепочечными или двухцепочечными ДНК или РНК. Гены вирусов могут быть заключены в одной хромосоме или разделены на несколько блоков (хромосом) , которые все вместе и составляют геном таких вирусов. Например, у ретвирусов геном представлен двухцепочечной РНК и состоит из десяти сегментов. Геномы вирусов, содержащих одноцепочечную РНК, также могут быть либо цельными (у ретровирусов ), либо сегментированными (у аренавирусов ). Геном РНК-содержащих вирусов представлен только линейными молекулами РНК. Все известные ДНК- содержащие вирусы позвоночных имеют геном, заключенный в одной хромосоме, линейной или кольцевой, одно- или двухцепочечной.Репликация геномов вирусов ДНК-геномы реплицируются клеточными или вирусоспецифическими ДНК-полимеразами. РНК-геномы реплицируются вирусоспецифическими РНК-полимеразами, которые также являются и транскриптазами. Репликация вирусных геномов происходит или одновременно с транскрипцией, или эти два процесса разделены во времени. Механизмы репликации геномов вирусов многообразны и определяются видом генома. Существует три модели репликации – полуконсервативная, консервативная и дисперсная. Консервативная и дисперсная модели репликации нуклеиновых кислот установлены только у вирусов. Полуконсервативная модель предполагает, что после первого раунда репликации одна цепь в каждой из двух дочерних молекул является родительской, другая – синтезируемой заново. По такой схеме реплицируются двунитевыеДНК-геномы вирусов. При реализации консервативной модели репликации одна дочерняя молекула состоит из двух родительских цепей, а другая – из вновь синтезированных цепей. Согласно консервативной модели реплицируются двунитевые РНК ротавирусов .ДНК-содержащие вирусы, реплицирующиеся таким образом, неизвестны. Дисперсная модель репликации приводит к образованию молекулы нуклеиновой кислоты, состоящей из фрагментов, как родительских цепей, так и вновь синтезированных.
Строение геномов эукариот и особенности их репликации
Геном эукариотДля клеток эукариот характерно наличие оформленного ядра. Информационной макромолекулой их генома является ДНК, которая неравномерно распределена по нескольким хромосомам в виде комплексов с многочисленными белками. Жизненно важная генетическая информация заключена и во внехромосомных молекулах ДНК. У эукариот — это ДНК хлоропластов, митохондрий и других пластид. Геном эукариот обладает избыточностью. Эукариотическая клетка содержит во много раз больше генов, чем прокариотическая. Большая часть их геномной ДНК представлена некодирующими последовательностями нуклеотидов. У эукариот около 5% ДНК составляют экзоны (участки гена, кодирующие белок), 25% – интроны (участки гена, которые транскрибируются, но затем удаляются при сплайсинге), а остальные 70% составляют спейсеры – нетранскрибируемые участки ДНК между генами.Особенности репликации у эукариот.Инициация репликации. Репликация начинается на определённом сайте молекулы ДНК, точке начала репликации или ori-сайте. В этой точке двойная спираль ДНК расплетается с образованием двух репликативных вилок, которые движутся по направлению друг к другу .В формировании репликативных вилок участвуют ДНК-топоизоимеразы и ДНК-хеликазы. По окончанию формирования репликативной вилки фермент ликвидирует разрыв и отделяется от ДНК.Разрыв водородных связей в двуцепочечной молекуле ДНК осуществляет фермент ДНК-хеликаза. Для этого фермент использует энергию макроэргических связей АТФ. В результате происходит раскручивание суперпсиральтной структуры ДНК в поддержании такой структуры участвуют SSB-белки. Эти белки не закрывают азотистых оснований и не препятствуют репликации.Элонгация – продолжение репликации. Репликация ДНК осуществляется ДНК-зависимыми ДНК-полимеразами.Субстартами для синтеза новой цепи ДНК являются дезоксинуклеозидтрифосфаты.. Эти же соединения служат источниками энергии. На образование каждой связи расходуются две макроэргические связи. В процессе также участвуют ионы магния, нейтрализуя отрицательный заряд нуклеотидов.У эукариот принимают участие 5 ДНК полимераз (α, β, γ, δ, ε), которые различаются по числу субъединиц, молекулярной массе, способности взаимодействовать с другими белками и функциональному назначению. Полимеразы β, δ и ε не могут начинать образование дочерней ДНК, т.к. не имеют сродства к одиночной цепи ДНК. Начинает репликацию полимераза α, которая синтезирует небольшой фрагмент РНК-праймер и далее примерно 60 нуклеотидных остатков ДНК. Образующийся небольшой двухцепочечный фрагмент позволяет присоединиться полимеразе δ и продолжить синтез новой цепи в направлении от 5/ к 3/. Выбор очередного нуклеотида определяется матрицей ДНК. При созревании отстающей цепи ДНК праймеры удаляют ДНК-полимераза β и присоединяет недостающие нуклеотиды. Восстановленные фрагменты соединяют ДНК-лигазы.
Вирусы - это внеклеточная форма жизни, обладающая собственным геномом и способная к воспроизведению только в клетках живых организмов.
Вирион (или вирусная частица) состоит из одной или нескольких молекул ДНК или РНК, заключенных в белковую оболочку (капсид), иногда содержащую также липидные и углеводные компоненты.
Вирусы размножаются только после инфицирования живых клеток. Различные вирусы проникают в животные и растительные клетки, а также бактерии (вирусы бактерий называются бактериофагами). Вирусы являются внутриклеточными паразитами на генетическом уровне и используют для своего размножения белок-синтезирующий аппарат клетки-хозяина.
По сравнению с геномами других организмов вирусный геном относительно мал и кодирует лишь ограниченное число белков, в основном белки капсида и один или несколько белков, участвующих в репликации и экспрессии вирусного генома. Необходимые метаболиты и энергия поставляются хозяйской клеткой.
Геном вирусов, заключенный внутри вирионов, может быть представлен ДНК или РНК, последние могут быть одно - и двуцепочечными, кольцевыми и линейными. Значения молекулярных масс ДНК вирусов находится в пределах 1∙10 6 - 200∙10 6 ; РНК - 1∙10 6 - 15∙10 6 дальтон.
По сложности строения генома вирусы широко варьируют - от фага Qβ (РНК-содержащий вирус бактерий), имеющего 4 гена, до вируса оспы (ДНК-содержащий вирус), геном которого насчитывает около 250 генов. Кроме того, все гены вируса могут быть заключены в одной молекуле нуклеиновой кислоты или распределены по нескольким молекулам, которые вместе и составляют геном такого вируса.
Типы генетического материала и механизм его репликации
У различных вирусов
3.1 РНК-содержащие вирусы (РНК → РНК)
Геномы почти всех известных РНК-содержащих вирусов, реплицирующихся по данной схеме - это линейные молекулы, их удобно разделить на 2 группы.
Первая группа -это однонитевые геномы положительнойполярности, т.е. с нуклеотидной последовательностью, соответствующей таковой у мРНК.

Такие геномы обозначают как (+) РНК. Цепь РНК такого типа может сразу после проникновения в клетку-хозяина обеспечивать синтез вирусных белков. Однонитевый (+) РНК-геном характерен для фага Qβ, вирусов табачной мозаики, полиомиелита, клещевого энцефалита. Вирусные (+) РНК-геномы кодируют несколько белков, среди которых РНК-зависимая РНК-полимераза (репликаза),способная синтезировать молекулы РНК без участия ДНК. С помощью этого фермента синтезируются сначала (-) нити РНК вируса, а затем репликаза осуществляет синтез (+) нити РНК. На заключительной стадии из накопившихся вирусных белков и (+) РНК формируются новые вирионы.
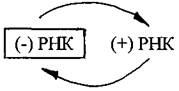
Этот вирусный фермент РНК-зависимая РНК-полимераза,синтезированная в предыдущем цикле размножения,упакован в вирионе в удобной для доставки в клетку форме. Инфекционный процесс начинается с того, что вирусный фермент копирует вирусный геном, образуя (+) РНК, которая выступает в качестве матрицы для синтеза вирусных белков, в том числе РНК-зависимой РНК-полимеразы, которая входит в состав образующихся вирионов.
ДНК-содержащие вирусы
Первая группа- вирусы с двуцепочечной ДНК, репликация которых осуществляется по схеме: ДНК→РНК→ДНК. Они получили название ретроидные вирусы. Представителями этой группы вирусов являются вирус гепатита В и вирус мозайки цветной капусты.
Репликация ДНК-генома этих вирусов осуществляется при посредстве промежуточных молекул РНК:
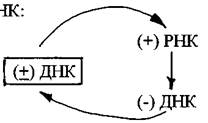
Молекулы (+) РНК образуются в результате транскрипциивирусных ДНК в клеточном ядре хозяйским ферментом ДНК-зависимой РНК-полимеразой.Транскрибируется только одна из нитей вирусной ДНК. Дальнейший синтез ДНК на РНК-матрице происходит в результате реакции, катализируемой обратной транскриптазой;сначала синтезируется (-) нить ДНК, а затем на вновь синтезированной (-) нити ДНК тот же фермент строит (+) нить. (+) РНК также используется для трансляции белков, необходимых для сборки вирусных частиц.
Вторая группа- вирусы с двухцепочечной ДНК, репликация которых осуществляется по схеме ДНК→ДНК. К этой группе относятся вирусы герпеса, оспы и др. В зараженной клетке ДНК-зависимая РНК-полимераза(фермент клетки-хозяина) транскрибирует с генома этих вирусов молекулы мРНК (т.е. (+) РНК), которые принимают участие в синтезе вирусных белков, а размножение вирусного генома осуществляет фермент ДНК-зависимая ДНК-полимераза:
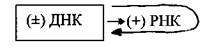
Бактериофаги
Отмеченные особенности характерны для вирусов животных, растений и бактерий. Последние, называемые бактериофагами, наиболее детально изучены с генетической точки зрения. Связано это, прежде всего с легкостью их культивирования в лабораторных условиях.
Разные фаги обнаруживают специфичность действия - они действуют на бактерии лишь определенного вида, редко на близкие виды и часто - на отдельные штаммы данного вида. Фаги со специфическим приспособлением к тому или иному хозяину отличаются от других фагов морфологией, размерами, биохимическими и серологическими свойствами.
Биохимический анализ выявляет в фаговой частице два компонента -белок и нуклеиновую кислоту. Белок образует хвост фага и мембрану головки. При резком изменении осмотической силы среды мембрана может разрываться и ДНК освобождается.
Практически все известные фаги содержат ДНК. Молекулы у них состоят из двух комплементарных нитей. Каждая фаговая частица содержит лишь одну молекулу ДНК.
Поскольку размножение фагов происходит внутри бактериальной клетки, рассмотрим типичную картину взаимодействия фага и чувствительной бактерии.
Фаговые частицы адсорбируются на поверхности бактерии, прикрепляясь к ее оболочке своими хвостами. Зараженная бактерия через 15-45 мин лизируется, освобождая 100 и более зрелых частиц следующего поколения фага. Встречая чувствительные бактериальные клетки, они вновь заражают их, и весь цикл повторяется. Если размножение фага происходит в культуре в жидкой среде, то оно приводит к просветлению (из-за лизиса клеток) суспензии бактерий. Размножение фага на бактериях, растущих на плотной среде, выявляется в виде зон лизиса среди сплошного газона бактерий. Эти пятна обнаженного агара называются негативными колониями фага.

Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого.
Поперечные профили набережных и береговой полосы: На городских территориях берегоукрепление проектируют с учетом технических и экономических требований, но особое значение придают эстетическим.

Папиллярные узоры пальцев рук - маркер спортивных способностей: дерматоглифические признаки формируются на 3-5 месяце беременности, не изменяются в течение жизни.
Читайте также:
