
Реферат морфология вирусов и грибов
Грибы — большая группа эукариотических организ-
мов, объединяющая свыше 100 000 видов. В систематике органи-
ческого мира грибы занимают особое положение. С животными
грибы сближает наличие в оболочке их клетки полисахаридной
субстанции — хитина (исключение составляют оомицеты, у кото-
рых обнаружена целлюлоза), участие в обмене азота — мочевины,
а в обмене углеводов — гликогена. Только в клетках животных
и грибов имеются цитохромы, участвующие в окислительно-вос-
становительных процессах. Однако по способу питания (адсорб-
тивное — путем всасывания, а не заглатывания пищи) и неогра-
ниченному росту грибы напоминают растения. Они не содержат
хлорофилла и по типу питания являются гетеротрофами: сапро-
фитические грибы используют остатки растительного или живот-
ного происхождения, а паразиты — ткани растений и животных
Проблемы систематики грибов до сих пор нельзя считать раз-
решенными. Рассматривается вопрос о выделении грибов в са-
мостоятельное царство, но в настоящее время пользуются бота-
нической классификацией. В ее основу положен морфологический
принцип строения тела грибов и органов полового воспроизве-
дения в культуре. Согласно этому грибы разделяются на выс-
щие и низшие, совершенные и несовершенные. Различают пять
классов:
фикомицеты (Phykomycetes) с тремя группами:
хитридиомицеты (Chitridiomycetes),
оомицеты (Oomycetes),
зигомицеты (Zygomycetes);
Архимицеты и фикомицеты относятся к низшим грибам. Пер-
вые из них мицелия не имеют, мицелий вторых несептирован. Эти
грибы обладают наиболее простой формой спороношения в виде
спорангиев, в которых заключены споры.
У аскомицетов и базидиомицетов, являющихся высшими гри-
бами, мицелий септированный (многоклеточный), строение ор-
ганов спороношения более сложное.
Представители первых четырех классов имеют половой и бес-
полый циклы развития и называются совершенными.
Дейтеромицеты являются высшими несовершенными гриба-
ми: они обладают септированным мицелием, но половое споро-
ношение у них отсутствует или не обнаружено. Группа несовер-
шенных грибов непостоянна, поскольку обнаружение у гриба
половых форм размножения позволяет отнести его к одному из
перечисленных классов.
Патогенные для человека грибы — возбудители микозов (от
греч. mykes — гриб) относят к различным систематическим груп-
пам ботанической классификации.
Грибы различают по величине, строению, местам произрас-
тания и физиологическим функциям. Их размеры варьируют от
микрометров (микроскопические грибы) до метров (шляпочные).
Исходя из особенностей питания и мест произрастания сформи-
рованы различные экологические группы: почвенные, фитопато-
генные, энтомофилы, зоофилы, антропофилы и др.
Наряду с различиями грибы имеют общие черты строения.
Грибы — это своеобразные эукариотические организмы, которые
в процессе своего развития претерпевают морфологические изме-
нения. Данные изменения могут быть связаны с жизненными
циклами — половым и бесполым или с влиянием факторов внеш-
ней среды. Основу клеточного строения большинства из них со-
ставляет масса тонких, ветвящихся трубчатых нитей, именуемых
гифами, а вся эта масса гиф называется мицелием (рис. 3). Диа-
метр гиф варьируется от 1 до 10 мкм, а их длина — от 4-6 до
80—100 мкм и более. Каждая гифа окружена тонкой жесткой стен-
кой, основным компонентом которой является хитин — азотсо-
держащий полисахарид, являющийся, как известно, структурным
 |
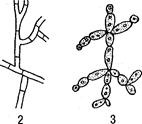 |
Рис. 3. Типы мицелия у грибов:
1 — несептированный (одноклеточный) мицелий; 2— септированный (многоклеточный) мицелий; 3 — псевдомицели и
компонентом наружного скелета членистоногих. Гифы не имеют клеточного строения. Протоплазма гиф либо совсем не разделена (у одноклеточных), либо разделяется поперечными перегородками, называемыми септами (у многоклеточных). Такие септы делят содержимое гиф на отдельные участки, внешне похожие на клетки, при этом образование септ не связано с делением ядер. В центре септы, как правило, остается небольшое отверстие (пора), через которое протоплазма может перетекать из одного компартмента в другой. В каждом компартменте могут находиться одно, два или несколько ядер. Гифы, не имеющие перегородок, образуют несептированный мицелий, а имеющие — септированный.
У дрожжевых и дрожжеподобных грибов образуется псевдомицелий, который в отличие от истинного мицелия, представляющего собой ветвистую трубку, разделенную у высших грибов поперечными перегородками, не имеет общей стенки, является цепочкой из клеток, которая формируется в процессе размножения почкованием.
Основой клеточной стенки грибов в отличие от бактерий являются полисахариды, состоящие из пяти-шести моносахаров. В связи с их меньшей, чем у бактериальных оболочечных белков, вариабельностью, наблюдается антигенное родство между далекими по своим морфологическим свойствам видами грибов. В состав полисахаридных фракций входят глюкозамин, манноза, глюкоза, ксилоза.
Непосредственно к внутренней части клеточной стенки прилегает цитоплазматическая мембрана, с которой в тесном контакте находится цитоплазматический ретикулюм, часто гранулярный, составляющий основную часть цитоплазмы. В ней расположены одно или несколько ядер, имеющих свою оболочку с порами, и ядрышко, содержащее в составе хромосом ДНК. В цитоплазме есть центральная вакуоль, а также митохондрии, микросомы, лизосомы, рибосомы, пластиды, комплекс Голъджи, глико-, липо-, хромопротеиды, секреторные гранулы, миелоидные образования, различные включения [волютин, гликоген, пигменты и др.).
Кроме того, в клетках могут накапливаться продукты метаболизма грибов — антибиотики, ферменты, витамины, токсины, и т. п.
При росте на питательной среде мицелий ветвится боковыми выростами гиф; переплетаясь, он образует грибницу. Различают мицелий вегетативный — погруженную в субстрат питающую часть грибницы; репродуктивный — воздушную, спороносящую часть грибницы; склероций — покоящийся тип грибов. Последний представляет собой округлое или продолговатое тело плотной консистенции диаметром от нескольких миллиметров до десятков сантиметров, образующееся веществами тесного переплетения гиф. Он богат запасными питательными веществами и обеспечивает жизнеспособность гриба в неблагоприятных условиях внешней среды (недостатке влаги, низких температурах и т. п.).
Различают два типа размножения грибов — половое и бесполое. По способу осуществления размножение может быть вегетативным, то есть происходить без образования специальных или или с помощью малодифференцированных органов размножения; репродуктивньш — путем образования специальных органов воспроизводства. Во втором случае возможно как бесполое, так и половое воспроизводство.
Вегетативное размножение грибов может осуществляться ку-сочками мицелия, склероциями, спорами, возникающими в результате расчленения гиф мицелия (хламидоспоры, артроспоры, бластоспоры, оидии, геммы).
Репродуктивное размножение грибов происходит при посредстве спор, возникающих половым или бесполым путем на специальных дифференцированных или спороносящих ветвях мицелия или органах.
Бесполое размножение осуществляется при помощи спор, развивающихся эндогенно (эндоспоры, спорангиеспоры) и экзогенно — на концах особых выростов мицелия или на мицелии (конидии).
Половое размножение у грибов осуществляется путем слияния мужских и женских половых гамет, в результате чего образуется зигота. Способы полового размножения у грибов весьма разнобразны. У низших грибов происходит слияние одинаковых (изогамия) или разных по размерам (гетерогамия) гамет. Из зиготы после периода покоя вырастает спорангиеносец со спорангием, наполненным эндоспорами. У высших грибов образуются для полового размножения аскоспоры и базидиоспоры.
У несовершенных грибов половой процесс заменяется гете-рокариозом (разноядерностью) и парасексуальным процессом |без слияния ядер в первом случае или со слиянием ядер во втором).
Патогенные для человека микроскопические грибы относятся к зигомицетам, аскомицетам, дейтеромицетам.
Зигомицеты — группа грибов класса фикомицетов; это назем-ные, чаще всего почвенные сапрофиты, мицелий в основном не-
септированный. Имеют особый тип полового процесса — слияние двух недифференцированных на гаметы клеток, с образованием покоящейся зигоспоры, с диплоидным набором хромосом. Бесполое размножение зигомицетов происходит с помощью эндоспор, образующихся в специальных крупных шаровидных клетках — спорангиях, которые формируются на свободных концах плодоносящих гиф — спорангиеносцах, имеющих разнообразную форму: грушевидную ( Mucor), шаровидную (Rhizopus), булавовидную (Асtinomucor) и др. Каждая спора дает начало новому мицелию.
Грибы родов Mucor, Rhizopus, Аbsidia, среди которых наиболее широко известен мукор, могут вызывать микозы у человека и животных, связанные с поражением кожи и легких.
Класс аскомицетов — одна из самых многочисленных и сравнительно высокоорганизованных групп микроскопических грибов, отличающаяся от зигомицетов сложностью строения, особенно репродуктивных органов. К ним относятся дрожжи и многие виды плесневых грибов, последние имеют расчлененный многоклеточный мицелий.
У аскомицетов в результате полового процесса, которому предшествует плазмогамия (слияние половых клеток), кариогамия (слияние ядер половых клеток) и мейоз, формируются специализированные клетки округлой формы — аски (сумки). В сумке образуется определенное число (чаще всего восемь) аскоспор, с помощью которых и происходит размножение. У одних аскомицетов сумки возникают непосредственно на мицелии, у других — внутри или на поверхности плодовых тел, образуемых в результате сплетения гиф мицелия.
Аспергиллы и пенициллы образуют сумки чаще всего округлой формы. У большинства аскомицетов плазмогамия и кариогамия при половом процессе разделены во времени. Бесполое размножение у дрожжей осуществляется почкованием, а у мицелиальных форм грибов — с помощью экзоспор (конидий), образующихся на конидиеносцах. Грибы рода Аspergillus и рода Реnicillium отличаются строением органов вегетативного спороношения. У аспергиллов на конидиеносцах находятся удлиненные клетки (стеригмы), которые формируют цепочки экзоспор — конидий. Плодоносящая гифа пенициллиума имеет три ряда стеригм, на которых располагаются округлые конидии в виде цепочек.
Среди аспергиллов и пенициллов встречаются антибиотико-образующие виды и патогенные для человека. Последние являются возбудителями таких микозов как аспергиллез и пенициллиоз (поражение кожи, слизистых, легких).
Дейтеромицеты — несовершенные грибы (Fungi imperfecti), объединяют более 25 000 видов, с септированным мицелием, весь их жизненный цикл проходит в гаплоидной стадии, без смены ядерных фаз. Мицелиальные грибы размножаются только с по-
мощью конидий. Из патогенных для человека к ним относятся возбудители дерматомикозов: трихофитии, микроспории, эпидер-мофитии, фавуса. Поражают кожу и ее придатки (волосы, ногти), вызывают тяжелые, склонные к хроническому протеканию заболевания, сопровождающиеся аллергией.
К несовершенным грибам относятся и дрожжеподобные грибы рода Саndidа, образующие в результате почкования псевдомицелия; у патогенных видов Саndidа присутствуют также покоящиеся формы в виде хламидоспор (толстостенных округлых, толненных липидами клеток, образовавшихся из вегетативных). Эти грибы являются условно-патогенными и могут вызывать поражение кожи, слизистых оболочек и внутренних органов (кандидозы или кандидомикозы). Поскольку грибы рода Саndidа являются представителями нормальной микрофлоры человека, поражение происходит чаще всего эндогенным путем на фоне снижения иммунной реактивности, дисбактериозов, нарушения обмена веществ, длительном применении антибактериальных средств широкого спектра действия.
Учебник предназначен для студентов, аспирантов и преподавателей высших медицинских учебных заведений, университетов, микробиологов всех специальностей и практических врачей.
5-е издание, исправленное и дополненное
Книга: Медицинская микробиология, иммунология и вирусология
Морфология грибов
К грибам относится большая группа одно– или многоклеточных организмов, которым присущи как признаки растений (неподвижность, неограниченный верхушечный рост, способность к синтезу витаминов, наличие клеточных стенок), так и животных (тип питания, наличие хитина в клеточных стенках, запасных углеводов в форме гликогена, образование мочевины, структура цитохромов). Различают грибы-паразиты, грибы-сапрофиты и грибы-симбионты. Грибы-паразиты поражают живые ткани растений, животных и человека, вызывая различные заболевания, называемые микозами. Из ныне известных свыше 100 000 видов грибов для человека патогенны не более 400. Грибы-сапрофиты питаются органическими веществами мертвых тканей или экскрементами. Примерами сапрофитов являются дрожжевые и плесневые грибы; последние также могут быть причиной микозов человека. Грибы-симбионты могут вступать в симбиотические отношения с корнями высших растений, в эту группу входят многие шляпочные грибы (базидиомицеты), в том числе и употребляемые в пищу, и ядовитые.
Грибы, паразитирующие в организме человека, вызывают микозы, протекающие с поражениями кожи, ее придатков или внутренних органов.
Единой общепринятой классификации грибов пока нет. Патогенные грибы, их роды и виды распределяются в различных семействах, подклассах и классах грибов: классе Deyteromycetes, или Fungi imperfecti (несовершенные грибы), классе Ascomycetes и классе Zygomycetes. Возбудитель криптококкоза относится к классу Basidiomycetes. В основе классификации грибов, которая более всего удобна для практики, лежат морфологические особенности и характер размножения.
Клетки грибов покрыты плотной клеточной оболочкой, состоящей из полисахаридов, близких к целлюлозе, и азотистых веществ, подобных хитину. У большинства грибов вегетативное тело (мицелий) состоит из системы тонких ветвящихся нитей, называемых гифами. Переплетаясь, мицелий образует грибницу. Гифы способны расти в длину и развиваются на поверхности или внутри питательного субстрата. Соответственно различают мицелий субстратный (вегетативный), врастающий в питательную среду, и воздушный. Концы нитей мицелия могут быть закручены в виде спиралей, завитков и т. д.
Грибы размножаются с помощью различных структур. При образовании половых спор имеет место мейоз, а конидии являются неполовыми репродуктивными органами. Половые стадии обнаружены у многих патогенных грибов, принадлежащих к классам Ascomycetes и Zygomycetes. У других патогенных грибов, которые относятся к дейтеромицетам, половые формы размножения не выявлены. У грибов, имеющих медицинское значение, существуют следующие разновидности половых спор:
а) зигоспоры – у некоторых зигомицетов верхушки расположенных близко друг к другу гиф сливаются, происходит мейоз, и образуются крупные зигоспоры с толстыми стенками (рис. 112, а);
б) аскоспоры – в специальных клетках, называемых асками (сумками), в которых произошел мейоз, образуется 4 – 8, иногда 16 и более спор, размеры, форма и поверхность которых могут быть весьма разнообразными у разных видов грибов (рис. 112, б);
в) базидиоспоры – после мейоза на поверхности особой клетки, называемой базидиумом, на вершине каждой из четырех стеригм развивается по одной круглой или слегка удлиненной базидиоспоре различных размеров (рис. 112, в).
У большинства грибов, имеющих медицинское значение, обнаруживаются разнообразные конидии (или экзоспоры), являющиеся формами неполового размножения. Если у гриба не известны половые стадии, классификация основывается на морфологических особенностях конидий. Они могут образовываться на специальных конидиофорах (конидиеносцах), по бокам или на концах обычных неспециализированных гиф или из гифальных клеток. В одной и той же колонии гриба может происходить образование нескольких типов конидий; в этом случае мелкие одноклеточные конидии называются микроконидиями, а крупные, часто многоклеточные, – макроконидиями.
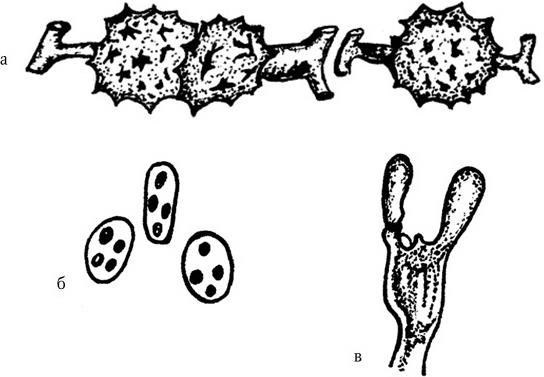
Рис. 112. Половые споры грибов:
а – зигоспоры; б – аскоспоры; в – базидиоспоры
Наиболее частыми типами конидий являются следующие виды спор:
а) бластоспоры – простые структуры, которые образуются в результате почкования с последующим отделением почки от родительской клетки, например у дрожжевых и дрожжеподобных грибов (рис. 113, а);
б) хламидоспоры – гифальные клетки увеличиваются, у них образуется толстая оболочка. Эти структуры высокоустойчивы к действию неблагоприятных факторов внешней среды и прорастают, когда условия становятся более благоприятными (рис. 113, б);
в) артроспоры – структуры, которые образуются в результате фрагментации гиф на отдельные клетки (рис. 113, в). Встречаются у дрожжеподобных грибов, возбудителя кокцидиоидоза, тканевых форм дерматофитов в волосе, кожных чешуйках и в ногте;
г) конидиоспоры – зрелые наружные споры – образуются на мицелии и не являются следствием превращения каких-либо других клеток грибницы. Они или возникают на дифференцированных конидиофорах, отличающихся от других нитей грибницы по размерам и форме (аспергилл, пеницилл, рис. 113, г, д), или располагаются по бокам и на концах любой ветви мицелия, прикрепляясь к ней непосредственно либо тонкой ножкой (рис. 113, е).

Рис. 113. Неполовое размножение грибов, морфология спор:
а – бластоспоры; б – интеркаларные (промежуточные) и терминальные (концевые) хламидоспоры; в – артроспоры; г – конидии аспергилла; д – конидии пеницилла; е – конидии споротрихума; ж – алейрии; з – спорангии с эндоспорами у мукора; и – сферулы кокцидиоидного гриба
Алейрии (рис. 113, ж) отличаются от обычных конидий тем, что при их образовании протоплазма соответствующих нитей целиком идет на формирование спор, от мицелия остаются нежизнеспособные фрагменты. С обилием алейрий связан мучнистый характер культур дерматофитов на плотных средах.
К эндоспорам совершенных грибов относятся спорангиоспоры мукоровых грибов; они развиваются в специальных органах, располагающихся на вершине спорангиеносца. Споры освобождаются при разрыве стенки спорангия и, попадая в благоприятные условия, прорастают и дают грибницы с соответствующими органами спороношения (рис. 113, з). Эндоспоры обнаруживаются также у тканевых форм возбудителей кокцидиоидоза и риноспоридиоза. Они развиваются в круглых образованиях – сферулах. В процессе созревания последних в них развиваются многочисленные округлые или яйцевидные тонкостенные эндоспоры (рис. 113, и). При разрыве стенки зрелой сферулы эндоспоры освобождаются и, попадая в благоприятные условия, повторяют жизненный цикл, постепенно превращаясь в зрелые сферулы.
Ни один из описанных выше морфологических элементов не является абсолютно характерным для того или иного вида гриба, так как в культурах разных грибов можно встретить одинаковые клеточные формы, различающиеся в деталях на разных этапах их созревания. Комплексом разнообразных клеточных элементов определяется большой полиморфизм грибов в культурах на различных питательных средах. В паразитарном состоянии тканевые формы многих грибов очень резко отличаются от культуральных. Морфологические черты тканевых форм более однообразны и более характерны для возбудителей соответствующих заболеваний.
ВНИМАНИЕ! САЙТ ЛЕКЦИИ.ОРГ проводит недельный опрос. ПРИМИТЕ УЧАСТИЕ. ВСЕГО 1 МИНУТА.
ТЕМА 2. МОРФОЛОГИЯ, ФИЗИОЛОГИЯ И КЛАССИФИКАЦИЯ ГРИБОВ, ПРОСТЕЙШИХ И ВИРУСОВ
1. Классификация, морфология и физиология грибов.
2. Классификация, морфология и физиология простейших.
3. Классификация, морфология и физиология вирусов.
Классификация, морфология и физиология грибов.
Общепринятой классификации грибов в настоящее время не существует, поэтому приведённые в литературе, сведения могут существенно различаться у разных авторов.
Грибы можно разделить на 7 классов:
По строению мицелия грибы подразделяются на группы:
1. низшие грибы (фикомицеты) – мицелий не имеет перегородок, к ним относятся классы:
- хитридиомицеты,- это водные грибы (сапрофитные или грибы, поражающие водоросли);
- гифохитридиомицеты – водные грибы (имеющие сходство с хитридиомицетами и оомицетами);
- оомицеты — паразиты высших растений и водяные плесени;
- зигомицеты - включают представителей рода Mucor, вызывающие мукоромикоз легких, головного мозга и других органов.
2.высшие грибы (эумицеты) – мицелий имеет перегородки (септы), к ним относятся:
- аскомицеты (сумчатые грибы) - представители родов Aspergillus (вызывающие аспергиллезы), Penicillium (вызывающие пенициллиозы) и дрожжи (вызывающие дрожжевые микозы);
- базидиомицеты (шляпочные грибы) –единственным патогенным для человека видом выступает Cryptococcus neoformans (вызывает криптококкоз — оппортунистический диссеминированный микоз, характеризующийся поражением ЦНС, реже лёгких, кожи, слизистых оболочек; редко развивается у людей с нормальным иммунитетом. Криптококкозный менингит — одно из наиболее характерных проявлений СПИДа у ВИЧ - инфицированных пациентов).
К базидиомицетам относятся и микроскопические грибы, и грибы с крупными плодовыми телами. Среди этих грибов есть паразиты растений (например, широко распространенные и очень опасные для сельскохозяйственных растений головневые и ржавчинные грибы), многочисленные почвенные сапрофиты — хорошо всем известные шляпочные грибы (например, шампиньоны, навозники). К базидиомицетам относятся и микоризообразующие шляпочные грибы, которые успешно развиваются только в тесном контакте с корнями древесных растений (например, белый, подберезовик, подосиновик и многие другие лесные грибы
- дейтеромицеты - дрожжеподобные грибы рода Candida, вызывающие у человека кандидоз (поражение кожи, слизистых оболочек и внутренних органов).
По способу размножения грибы разделяют на:
1. Совершенные, имеющие и половой и бесполый способ размножения, к ним относятся зигомицеты, аскомицеты и базидиомицеты.
2. Несовершенные, размножающиеся только бесполым путем – дейтеромицеты.
Морфология и физиология грибов.
Грибы – эукариоты, имеющие чётко ограниченное ядро. По типу питания они гетеротрофы (то есть используют в качестве источника энергии и углерода органические соединения). По типу дыхания – аэробы. Грибы имеют ядро с ядерной оболочкой, цитоплазму с органеллами, цитоплазматическую мембрану и толстую клеточную стенку. Грибы имеют некоторые сходства и с растениями и с животными.
Признаки, придающие определённые сходства с растениями:
- наличие клеточной стенки;
- способность к неограниченному росту;
- необходимость прикрепления к субстрату;
- неподвижность в вегетативном состоянии;
- способ размножения и распространение спорами.
Признаки, придающие определённые сходства с животными:
- гетеротрофный тип питания;
- потребность в витаминах;
- способность к синтезу хитина;
- образование и накопление мочевины и гликогена (а не крахмала).
Различают плесневые и дрожжевые формы грибов.
Плесневые(гифальные) грибы образуют ветвящиеся тонкие нити (гифы), сплетающиеся в мицелий (плесень). Гифы, врастающие в питательный субстрат, называются вегетативными гифами (отвечают за питание гриба), а растущие над поверхностью субстрата — воздушными или репродуктивными гифами (отвечают за бесполое размножение).
Дрожжи и дрожжеподобныегрибы представлены отдельными овальными клетками, морфологически сходными между собой.
У грибов выделяют половой и бесполый (вегетативный) типы размножения.
При бесполом размножении возможны процессы почкования (характерны для дрожжеподобных грибов) и спорообразования. Дочерние клетки, образующиеся при почковании дрожжей, называют бластоспорами.
Среди спор бесполого размножения различают экзо- и эндоспоры. Экзоспоры образуются открыто на вершине гиф снаружи их. Такие споры называются конидиями, а гифы, несущие их, конидиеносцами.
Эндоспоры (спорангиоспоры) образуются в особых образованиях - спорангиях, а гифы, несущие их - спорангиеносцами
Среди спор полового размножения грибов различают: аскоспоры (образуют аскомицеты), базидиоспоры (образуют базидиомицеты), зигоспоры (образуют зигомицеты).
Основные уровни клеточной организации живых систем. Общая характеристика, формы и размеры бактерий, строение бактериальной клетки. Характеристика и отличительные особенности грибов. Дрожжи: характеристика, процесс размножения, форма и строение клетки.
Нажав на кнопку "Скачать архив", вы скачаете нужный вам файл совершенно бесплатно.
Перед скачиванием данного файла вспомните о тех хороших рефератах, контрольных, курсовых, дипломных работах, статьях и других документах, которые лежат невостребованными в вашем компьютере. Это ваш труд, он должен участвовать в развитии общества и приносить пользу людям. Найдите эти работы и отправьте в базу знаний.
Мы и все студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будем вам очень благодарны.
Чтобы скачать архив с документом, в поле, расположенное ниже, впишите пятизначное число и нажмите кнопку "Скачать архив"
| Рубрика | Биология и естествознание |
| Вид | реферат |
| Язык | русский |
| Дата добавления | 20.03.2016 |
| Размер файла | 339,9 K |
История микроскопа и изучение морфологии микроорганизмов как собирательной группы живых организмов: бактерии, археи, грибы, протисты. Формы, размер, морфология и строение бактерий, их классификация и химический состав. Строение и классификация грибов.
реферат [130,0 K], добавлен 05.12.2010
Исследование морфологических признаков бактерий, микроскопических грибов и дрожжей. Изучение внешнего вида, формы, особенностей строения, способности к движению, спорообразованию, способов размножения микроорганизмов. Форма и строение дрожжевой клетки.
реферат [28,8 K], добавлен 05.03.2016
Систематика - распределение микроорганизмов в соответствии с их происхождением и биологическим сходством. Морфология бактерий, особенности строения бактериальной клетки. Морфологическая характеристика грибов, актиномицетов (лучистых грибов) и простейших.
реферат [27,2 K], добавлен 21.01.2010
Споры – форма бактерий с грамположительным типом строения клеточной стенки. Роль спорообразования бактерий и грибов для практики. Строение и особенности химического состава бактериальной споры. Микробиологическое обоснование пастеризации и стерилизации.
контрольная работа [223,5 K], добавлен 02.10.2011
Систематика микроорганизмов по фенотипическим, генотипическим и филогенетическим признакам. Отличия прокариот и эукариот, анатомия бактериальной клетки. Морфология микроорганизмов: кокки, палочки, извитые и нитевидные формы. Генетическая система бактерий.
презентация [6,4 M], добавлен 13.09.2015
Видоизменения мицелия в процессе приспособления к различным наземным условиям обитания. Размножение, питание и классификация грибов, их значение в биосфере и народном хозяйстве. Строение клетки гриба и бактериальной клетки, жизнедеятельность грибов.
реферат [198,1 K], добавлен 05.06.2010
Особенности питания и строения грибов как отдельной группы микроорганизмов. Рост гифов гриба и строение клеточной стенки гифа (липиды, хитин). Характеристика способов размножения грибов: вегетативное, почкообразование, спорообразование, деление клетки.
презентация [665,0 K], добавлен 25.02.2015
История открытия микроорганизмов. Клеточная стенка — структурный элемент бактериальной клетки, ее строение у грамотрицательных и грамположительных бактерий. Состав гомогенного слоя клеточной стенки. Функция пептидогликана; периплазматическое пространство.
реферат [1,8 M], добавлен 15.05.2012
Формы и размеры бактериальных организмов и их краткая характеристика. Строение бактериальной клетки, движение бактерий. Спорообразование и его биологическая роль, размножение бактерий. Передача признаков с помощью процессов трансдукции и трансформации.
лекция [25,5 K], добавлен 25.03.2013
Группа микроскопических одноклеточных организмов-прокариотов. Микроскопические методы исследования микроорганизмов. Формы, строение и химический состав бактериальной клетки. Функции поверхностных структур. Дыхание, питание, рост и размножение бактерий.
презентация [3,8 M], добавлен 24.01.2017
Автор работы: Пользователь скрыл имя, 15 Декабря 2014 в 19:00, реферат
12 февраля 1892 г. Д.И.Ивановский открыл вирусы - новую форму существования жизни. Своими исследованиями он заложил основы ряда научных направлений вирусологии: изучение природы вируса, цитопаталогических вирусных инфекций, фильтрующихся форм микроорганизмов, хронического и латентного вирусоносительства. Один из выдающихся советских фитовирусологов В.Л.Рыжков писал: “Заслуги Д.И.Ивановского не только в том, что он открыл совершенно новый вид заболеваний, но и в том, что он дал методы их изучения”.
1. История
2. Строение вирусов
3. Основные методы культивирования вирусов
4. Взаимодействие вируса с клеткой хозяина
5. Возбудители гриппа
6. Вирусы ECHO
7. Вирусы Коксаки
8. Заключение
9. Литература
морфология вирусов.docx
МИНИСТЕРСТВО ЗДРАВООХРАНЕНИЯ ОМСКОЙ ОБЛАСТИ
Бюджетное образовательное учреждение Омской области
Выполнили: Чернявский Д.Д.
студенты Гр. М-27 Подгруппы: А
Омск 2014
Содержание:
- История
- Строение вирусов
- Основные методы культивирования вирусов
- Взаимодействие вируса с клеткой хозяина
- Возбудители гриппа
- Вирусы ECHO
- Вирусы Коксаки
- Заключение
- Литература
1.История
Заболевания растений, животных и человека, вирусная природа которых в настоящее время установлена, в течении многих столетий наносили ущерб хозяйству и вред здоровью человека. Хотя многие из этих болезней были описаны, но попытки установить их причину и обнаружить возбудитель оставались безуспешными. В 80-е годы XIX века – Луи Пастер впервые применил термин вирус (от лат. virus – яд) для обозначения инфекционного агента. В результате наблюдений Д.И.Ивановский и В.В.Половцев впервые высказали предположение, что болезнь табака, описанная в 1886 году A.D.Mayer в Голландии под название мозаичной, представляет собой не одно, а два совершенно различных заболевания одного и того же растения: одно из них - рябуха, возбудителем которого является грибок, а другое неизвестного происхождения. Исследование мозаичной болезни табака Д.И.Ивановский продолжает в Никитинском ботаническом саду (под Ялтой) и ботанической лаборатории Академии наук и приходит к выводу, что мозаичная болезнь табака вызывается бактериями, проходящими через фильтры Шамберлана, которые, однако, не способны расти на искусственных субстратах. Возбудитель мозаичной болезни называется Ивановским то “фильтрующимися” бактериями, то микроорганизмами, так как сформулировать сразу существование особого мира вирусов было весьма трудно. Подчеркивая, что возбудитель мозаичной болезни табака не мог быть обнаружен в тканях больных растений с помощью микроскопа и не культивировался на искусственных питательных средах. 12 февраля 1892 г. Д.И.Ивановский открыл вирусы - новую форму существования жизни. Своими исследованиями он заложил основы ряда научных направлений вирусологии: изучение природы вируса, цитопаталогических вирусных инфекций, фильтрующихся форм микроорганизмов, хронического и латентного вирусоносительства. Один из выдающихся советских фитовирусологов В.Л.Рыжков писал: “Заслуги Д.И.Ивановского не только в том, что он открыл совершенно новый вид заболеваний, но и в том, что он дал методы их изучения”. 1898 – Фредерик Лёффлер и Пауль Фрош выделили первый вирус животных – вирус ящура. В 1898 г. немецкие учёные Лёфлер и Форш открыли вирусную болезнь животных - ящур рогатого скота. В 1935 году У. Стенли из сока табака, пораженного мозаичной болезнью, выделил в кристаллическом виде ВТМ (вирус табачной мозаики). За это в 1946 году ему была вручена Нобелевская премия. В 1958 году Р.Франклин и К.Холм, исследуя строение ВТМ, открыли, что ВТМ является полым цилиндрическим образованием. В 1960 году Гордон и Смит установили, что некоторые растения заражаются свободной нуклеиновой кислотой ВТМ, а не целой частицей нуклеотида. В этом же году крупный советский ученый Л.А.Зильбер сформулировал основные положения вирусогенетической теории. В 1962 году американские ученые А.Зигель, М.Цейтлин и О.И.Зегал экспериментально получили вариант ВТМ, не обладающий белковой оболочкой, выяснили, что у дефектных ВТМ частиц белки располагаются беспорядочно, и нуклеиновая кислота ведет себя, как полноценный вирус. В 1968 году Р.Шепард обнаружил ДНК-содержащий вирус. Одним из крупнейших открытий в вирусологии является открытие американских ученых Д.Балтимора и Н.Темина, которые нашли в структуре ретровируса ген, кодирующий фермент - обратную транскриптазу. Назначение этого фермента - катализировать синтез молекул ДНК на матрице молекулы РНК. За это открытие они получили Нобелевскую премию. В знак признания выдающихся заслуг Д.И.Ивановского перед вирусологической наукой Институту вирусологии АМН СССР в 1950 году было присвоено его имя, в Академии медицинских наук учреждена премия имени Д.И.Ивановского, присуждаемая один раз в три года. По мере изучения природы вирусов в первом полстолетия после их открытия Д.И.Ивановским (1892) формировались представления о вирусах как о мельчайших организмах. Наиболее правдоподобной и приемлемой является гипотеза о том, что вирусы произошли из “беглой” нуклеиновой кислоты, т.е. нуклеиновой кислоты, которая приобрела способность реплицироваться независимо от той клетки, из которой она возникла, хотя при этом предусматривается, что такая ДНК реплицируется с использованием структур этой или другой клеток.
2.Строение вирусов
Вирус (от лат. virus — яд) — простейшая форма жизни, микроскопическая частица, представляющая собой молекулы нуклеиновых кислот (ДНК или РНК), заключённые в белковую оболочку и способные инфицировать живые организмы. Вирусы являются облигатными паразитами, так как не способны размножаться вне клетки. В настоящее время известны вирусы, размножающиеся в клетках растений, животных, грибов и бактерий (последних обычно называют бактериофагами). Обнаружены также вирусы, поражающие другие вирусные частицы (вирусы-сателлиты). Вирусы имеют генетические связи с представителями флоры и фауны Земли. Согласно последним исследованиям, геном человека более чем на 32 % состоит из информации, кодируемой вирусоподобными элементами. С помощью их происходит горизонтальный перенос генов, то есть передача генетической информации не от непосредственных родителей к своему потомству, а между двумя неродственными (или даже относящимися к разным видам) особями.
Отличительные признаки вирусов:
- содержат лишь один тип нуклеиновой кислоты (РНК или ДНК);
- не имеют собственных белоксинтезирующих и энергетических систем;
- не имеют клеточной организации;
- обладают разобщенным способом репродукции (синтез белков и нуклеиновых кислот происходит в разных местах и в разное время);
- облигатный паразитизм вирусов реализуется на генетическом уровне;
- вирусы проходят через бактериальные фильтры.
Вирусные частицы могут существовать в двух формах: внеклеточной (вириона) и внутриклеточной (вируса). По форме бывают: шаровидные (вирус гриппа); кубоидальные (оспы); палочковидные (мозаичная болезнь табака, картофеля); сперматозоидные (многие бактериофаги); пулевидные (вирус бешенства); нитевидные (колифаги). Размеры их колеблются от 15 - 18 до 300 - 400 нм. В центре вириона — вирусная нуклеиновая кислота, покрытая белковой оболочкой — капсидом, который состоит из капсомеров.
- Защита генетического материала (ДНК или РНК) вируса от механических и химических повреждений.
- Определение потенциала к заражению клетки.
- На начальных стадиях заражения клетки: прикрепление к клеточной мембране, разрыв мембраны и внедрение в клетку генетического материала вируса.
Нуклеиновая кислота и капсидная оболочка составляют нуклеокапсид. Нуклеокапсид сложноорганизованных вирионов покрыт внешней оболочкой – суперкапсидом. Суперкапсид (дополнительная липопротеидная) оболочка образована из плазматической мембраны клетки-хозяина. Она встречается только у сравнительно больших вирусов (грипп, герпес). Эта наружная оболочка является фрагментом ядерной или цитоплазматической мембраны клетки-хозяина, из которой вирус выходит во внеклеточную среду. Иногда в наружных оболочках сложных вирусов помимо белков содержатся углеводы, например у возбудителей гриппа и герпеса.
Строение ДНК- и РНК- содержавших вирусов принципиально не отличается от нуклеиновой кислоты других микроорганизмов.
- двухцепочечной;
- одноцепочечной;
- кольцевой;
- двухцепочечной, но с одной более короткой цепью;
5. двухцепочечной, но с одной непрерывной, а с другой фрагментированной цепями.
- однонитевой;
- линейной двухнитевой;
- линейной фрагментированной;
- кольцевой; содержащей две одинаковые однонитевые РНК.
Вирусные белки подразделяют на:
- геномные — нуклеопротеиды. Обеспечивают репликацию вирусных нуклеиновых кислот и процессы репродукции вируса. Это ферменты, за счет которых происходит увеличение количества копий материнской молекулы, или белки, с помощью которых на матрице нуклеиновой кислоты синтезируются молекулы, обеспечивающие реализацию генетической информации;
- белки капсидной оболочки — простые белки, обладающие способностью к самосборке. Они складываются в геометрически правильные структуры, в которых различают несколько типов симметрии: спиральный, кубический (образуют правильные многоугольники, число граней строго постоянно) или смешанный;
- белки суперкапсидной оболочки — это сложные белки, разнообразные по функции. За счет них происходит взаимодействие вирусов с чувствительной клеткой. Среди белков суперкапсидной оболочки выделяют:
а) якорные белки (одним концом они располагаются на поверхности, а другим уходят в глубину; обеспечивая контакт вириона с клеткой);
б) ферменты (могут разрушать мембраны клеток);
в) гемагглютинины (вызывают гемагглютинацию);
г) элементы клетки хозяина.
Особенность строения вирусного генома заключается в том, что в зависимости от типа вируса наследственная информация записывается либо на ДНК, либо на РНК. Мутации у вирусов могут возникать спонтанно, в процессе репликации нуклеиновой кислоты вируса или под влиянием тех же внешних факторов, мутагенов, что и у бактерий. Фенотипические мутации вирусного генома проявляются изменениями в антигенной структуре, неспособностью вызывать продуктивную инфекцию в клетке, зависимостью репродукции от температуры. Свойства вирусов изменяются при одновременном заражении несколькими вирусами чувствительной клетки.
3.Основные методы культивирования вирусов
Биологический - заражение лабораторных животных. После заражения вирусами лабораторных животных происходит их гибель. По изменениям в тканях после вскрытия судят о той или иной вирусной инфекции. Культивирование вирусов в развивающихся куриных эмбрионах. Куриные эмбрионы выращивают в инкубаторе 7 - 10 дней, а затем используют для культивирования. В результате заражения происходит гибель эмбриона. При микрокопировании препарата - появления бляшек (скопления погибших клеток с вирионами). Размножение в культуре ткани (это основной метод культивирования вирусов). Для поддержания клеток культуры ткани используют специальные среды. Это жидкие питательные среды сложного состава, содержащие аминокислоты, углеводы, факторы роста, источники белка, антибиотики и индикаторы для оценки развития клеток культуры ткани. О репродукции вирусов в культуре ткани судят по их цитопатическому действию, которое носит разный характер в зависимости от вида вируса.
Основные проявления цитопатического действия вирусов:
- размножение вируса может сопровождаться гибелью клеток или морфологическими изменениями в них;
- некоторые вирусы вызывают слияние клеток и образование многоядерного синцития;
- клетки могут расти, но делиться, в результате чего образуются гигантские клетки;
- в клетках появляются включения (ядерные, цитоплазматические, смешанные). Включения могут окрашиваться в розовый цвет (эозинофильные включения) или в голубой (базофильные включения);
- если в культуре ткани размножаются вирусы, имеющие гемагглютинины, то в процессе размножения клетка приобретает способность адсорбировать эритроциты (гемадсорбция).
4.Взаимодействие вируса с клеткой хозяина
Существует четыре типа взаимодействия:
- продуктивная вирусная инфекция (взаимодействие, в результате которого происходит репродукция вируса, а клетки погибают);
- абортивная вирусная инфекция (взаимодействие, при котором репродукции вируса не происходит, а клетка восстанавливает нарушенную функцию);
- латентная вирусная инфекция (идет репродукция вируса, а клетка сохраняет свою функциональную активность);
- вирусиндуцированная трансформация (взаимодействие, при котором клетка, инфицированная вирусом, приобретает новые, ранее не присущие ей свойства).
Читайте также:
