
Обратная транскрипция вирусная трансдукция и их значение для генетической инженерии
Обратная транскрипция — создание ДНК-копии по матрице РНК
Это правило переноса генетической информации составляет суть молекулярной биологии. Только один раз, в 1970 г., оно было модифицировано, когда признали существование обратной транскрипции. Впервые обратная транскрипция была обнаружена у опухолевых вирусов мышей и кур. Сейчас эти РНК-сод ержашие вирусы называют ретровирусами. Их инфекционный цикл хорошо изучен. Вирус проникает в клетку-мишень, и на основе своей РНК создает копию ДНК, которая встраивается в хромосому хозяина. При делении клетки встроенная копия ДНК вирусного генома удваивается и передается дочерним клеткам. Таким образом, наследственный материал вируса оказывается включенным в геном клетки. Вирусная РНК может образоваться позже путем копирования встроенной ДНК. Это приведет к образованию новых инфекционных вирусов.
Именно таким образом вирус иммунодефицита человека (ВИЧ) встраивается в ядерную ДНК и размножается вместе с клеточным геномом. Вот почему от этого вируса трудно избавиться, ведь каждая встроенная (интегрированная) ДНК-копия вируса может использоваться для образования множества копий РНК в результате нормального процесса транскрипции, а это в свою очередь приводит к появлению новых инфекционных ВИЧ-частиц. Еще один пример ретровирусов — это вирусы, вызывающие рак легких и лейкоз у человека и животных.
Похожие главы из других книг:
Глава 6 ОБРАТНАЯ СВЯЗЬ СОМЫ И ЗАРОДЫШЕВОЙ ЛИНИИ В этой главе мы рассматриваем, как могли изменяться последовательности ДНК вариабельных генов антител в клетках зародышевой линии за эволюционное время. Некоторые данные указывают на существование обратной связи генов, т.
Перезапись генетического текста и перевод в белковый текст (транскрипция и трансляция) В клетке ДНК служит в качестве матрицы, на которой первоначально происходит синтез разных РНК. Процесс перезаписи генетической информации из ДНК в РНКовый текст получил название
Перезапись генетического текста и перевод в белковый текст (транскрипция и трансляция) В клетке ДНК служит в качестве матрицы, на которой первоначально происходит синтез разных РНК. Процесс перезаписи генетической информации из ДНК в РНКовый текст получил название
Целенаправленное создание новых генов В некоторых случаях сомневаться в способности клетки направлять мутационный процесс на нужные гены не приходится. Особенно ярко проявляется это в работе иммунной системы позвоночных. Биологов и медиков давно интересовал вопрос,
7. БИОЛОГИЧЕСКАЯ ОБРАТНАЯ СВЯЗЬ (БОС) Интерес к исследованиям биологической обратной связи (БОС) в целях произвольного управления ФС на основе объективной информации о динамике психофизиологических показателей возник в связи с данными об изменениях в ФС человека,
Обратная транскрипция Некоторые РНК-содержащие вирусы (вирус саркомы Рауса, ВИЧ) обладают уникальным ферментом – РНК-зависимой ДНК-полимеразой, часто называемой обратной транскриптазой или ревертазой. Этот фермент обладает время активностями. Первая из них –
Транскрипция Транскрипцией называется процесс переноса генетической информации с ДНК на РНК. Матрицей для синтеза РНК служит только одна из двух нитей ДНК (так называемая смысловая цепь). Транскрипция происходит не на всей молекуле ДНК, а на участке одного гена. Ген –
Рокфеллер финансирует создание агробизнеса В то время когда братья Рокфеллеры с помощью своего плана Зеленой революции расширяли сферу влияния своего глобального бизнеса в развивающихся странах от нефти до сельского хозяйства, они финансировали малозаметный проект в
Создание орудий мышления Как нельзя голыми руками заниматься плотницким делом, так и мышление редко может осуществляться с помощью голого мозга Бо Дальбом, Ларс-Эрик Джанлерт. Компьютерное будущее (в печати). Перед каждым агентом стоит задача наилучшего использования
Взаимодействие эндокринных систем: прямая связь, обратная связь, синергизм, пермиссивное действие, антагонизм Как уже было упомянуто, отдельные эндокринные железы и их гормоны взаимодействуют друг с другом, обеспечивая функциональное единство организма. Между
Теория главной копии По мнению ведущего молекулярного биолога Харриса Бернштейна (Harris Bernstein) из Аризонского университета, половой процесс возник для починки генов. Первым намеком на это стало исследование особых мутантных дрозофил, не способных репарировать
Создание Олиновского нейропсихиатрического исследовательского центра Одним из главных спонсоров Института жизни было семейство Олин, у которого несколько детей лечились в ИЖ. Доктор Хэнк Шварц решил назвать наше новое здание в тюдоровском стиле в их честь. Теперь
Создание каталога мира: вербально-концептуальная ассоциативная зона Вербально-концептуальная ассоциативная зона, расположенная в месте пересечения височной, теменной и затылочной долей, в первую очередь отвечает за формирование абстрактных концепций и за их
При изучении ретровирусов, геном которых представлен молекулами одноцепочечной РНК, было обнаружено, что в процессе внутриклеточного развития ретровирус проходит стадию интеграции своего генома в виде двухцепочечной ДНК в хромосомы клетки-хозяина. В 1964 г. Темин выдвинул гипотезу о существовании вирусспецифичного фермента, способного синтезировать на РНК-матрице комплементарную ДНК. Усилия, направленные на выделение такого фермента, увенчались успехом, и в 1970 г. Темин с Мизутани, а также независимо от них Балтимор открыли искомый фермент в препарате внеклеточных вирионов вируса саркомы Рауса. Данная РНК-зависимая ДНК-полимераза получила название обратная транскриптаза, или ревертаза.
Каждый вирион (полноценная вирусная частица, состоящая из нуклеиновой кислоты и белковой оболочки) ретровирусов содержит две идентичные цепи РНК размером от 8000 до 10 000 нуклеотидов. Области 5'- и 3'-концов обеих цепей модифицированы, как у всех эукариотических мРНК (5'-кэпы, З'-полиадениловые хвосты). Вирусные РНК имеют 5 структурных элементов: 1) прямые повторы на 5'- и З'-концах РНК (R); 2) последовательность из 80 -- 120 нуклеотидов, находящуюся около 5 концевого повтора (U5); 3) последовательность из 170 -- 1200 нуклеотидов около З'-концевого повтора (U3); 4) последовательность из 15 20 нуклеотидов (Р), в пределах которой клеточная тРНК комплементарно взаимодействует с ретровирусной РНК, что создает праймер для синтеза первой цепи ДНК; 5) сегмент Pu, находящийся непосредственно перед повтором U3 и являющийся сайтом для праймирования второй цепи ДНК -- такой сегмент одинаков у РНК всех ретровирусов определенного типа.
Этапы обратной транскрипции:
5. К моменту завершения синтеза первой цепи ДНК большая часть вирусной РНК разрушается РНКазой Н.
6. Синтез З'-конца второй цепи ДНК.
7. Удаление тРНК и оставшегося участка вирусной (+) РНК РНКазой Н.
8. Второй прыжок, в результате которого новосинтезированная вторая цепь ДНК комплементарно взаимодействует с тРНК-связывающей последовательностью первой цепи.
9. Удлинение З'-концов каждой цепи, образование дуплекса ДНК.
Вся последовательность реакций протекает без явного участия ферментов репликации клетки-хозяина (топоизомеразы, хеликазы, праймазы, ДНК-связывающего белка, лигазы и т.д.). При этом следует отметить, что молекулы вирусных ДНК длиннее молекул вирусных РНК, которые послужили матрицей для обратной транскрипции. Действительно, к 5'-концу (+) цепи вирусной ДНК добавилась последовательность U3, а к 3 -концу этой цепи -- последовательность U5. В результате на концах молекулы вирус специфической ДНК появился длинный (несколько сотен нуклеотидов) концевой повтор (ДКП или LTR.), имеющий структуру U3U5.

Рис.2. Схема обратной транскрипции ретровирусной РНК с образованием двуцепочечной ДНК (Darnell J., et.al. Molecular Cell Biology. - N. Y.: Scientific Amer. Books, 1986. - P. 1052)
Синтез ДНК на РНК-матрице in vitro и ревертаза используется в генетической инженерии для синтеза генов и их фрагментов, а также целенаправленного синтеза на матричных РНК комплементарных молекул ДНК (кДНК) для расшифровки первичной структуры РНК и белков.

Рис. 3. Схема получения кДНК с использованием ревертазы вируса и трех дополнительных ферментов: поли (А)-полимеразы, фрагмента Кленова ДНК-полимеразы I и нуклеазы S1. (Цитировано по)
Реакцию обратной транскрипции проводят в специально подобранных условиях с использованием сильных ингибиторов РНКазной активности. При этом удается получать полноразмерные ДНК-копии целевых молекул РНК. В качестве праймера при обратной транскрипции поли (А) - содержащих мРНК используют олигo (dT)-праймер, а для молекул РНК, не имеющих З'-поли (А) концов, -- химически синтезированные олигонуклеотиды, комплементарные З'-концу изучаемой РНК. После синтеза на мРНК комплементарной цепи ДНК и разрушения РНК (обычно применяют обработку щелочью) осуществляют синтез второй цепи ДНК. При этом используют способность ревертазы образовывать на 3'-концах одноцепочечных кДНК самокомплементарные шпильки, которые могут выполнять функции праймера. Матрицей служит первая цепь кДНК. Данная реакция может катализироваться как ревертазой, так и ДНК-полимеразой I E. coli. Показано, что сочетание этих двух ферментов позволяет повысить выход полноценных двухцепочечных молекул кДНК. По окончании синтеза первая и вторая цепи кДНК остаются ковалентно связанными петлей шпильки, служившей праймером при синтезе второй цепи. Эту петлю расщепляют эндонуклеазой S1, специфически разрушающей одноцепочечные участки нуклеиновых кислот. Образующиеся при этом концы не всегда оказываются тупыми, и для повышения эффективности последующего клонирования их репарируют до тупых с помощью фрагмента Кленова ДНК-полимеразы I E. сoli (остающаяся часть молекулы, которая сохраняет присущие ей каталитические активности).
Уже одно это открытие формально поставило РНК в центр основного постулата молекулярной генетики, так как показало, что поток генетической информации распространяется от РНК не в одном, а в двух направлениях: не только к белку, но и к ДНК. Все более глубокое проникновение в механизмы основных молекулярно-генетических процессов (репликацию, транскрипцию и трансляцию) способствовало возникновению понятия о неканонических функциях РНК, осознанию полифункциональности рибонуклеиновых кислот.
Современная биология поражает уникальностью и масштабностью своих открытий. На сегодняшний день эта наука изучает большинство процессов, которые скрыты от нашего глаза. Этим примечательна молекулярная биология – одно из перспективных направлений, которое помогает разгадать сложнейшие тайны живой материи.
Что такое обратная транскрипция
Обратная транскрипция (сокращенно ОТ) – это специфический процесс, характерный для большинства РНК-содержащих вирусов. Главной его особенностью является синтез двухцепочечной молекулы ДНК на базе матричной РНК.
ОТ не характерна для бактерий или эукариотических организмов. Главный фермент – ревертаза – играет ключевую роль в синтезе двухцепочечной ДНК.
История открытия
Идея о том, что молекула рибонуклеиновой кислоты может стать матрицей для синтеза ДНК, считалась абсурдной вплоть до 1970 года. Тогда Балтимор и Темин, работавшие отдельно друг от друга, почти одновременно открыли новый фермент. Они назвали его РНК-зависимая-ДНК-полимераза, или обратная транскриптаза.
Открытие данного энзима безоговорочно подтвердило существование организмов, способных к обратной транскрипции. В 1975 году оба ученых получили Нобелевскую премию. Через некоторое время Энгельгардт предложил альтернативное название обратной транскриптазы – ревертаза.

Почему ОТ противоречит центральной догме молекулярной биологии
Центральная догма – это принципиальная схема последовательного синтеза белка в любой живой клетке. Такая схема строится из трех компонентов: ДНК, РНК и белок.
Согласно центральной догме, РНК может синтезироваться исключительно на матрице ДНК, а уж затем РНК участвует в построении первичной структуры белка.
Эта догма была официально принята в научном обществе раньше, чем произошло открытие обратной транскрипции. Неудивительно, что идея об обратном синтезе ДНК из РНК долго отвергалась учеными. Лишь в 1970 году вместе с открытием ревертазы была поставлена точка в этом вопросе, что отразилось и в принципиальной схеме синтеза белка.
Ревертаза ретровирусов птиц
Процесс обратной транскрипции не обходится без участия РНК-зависимой-ДНК-полимеразы. Максимально изучена на сегодняшний день ревертаза ретровируса птиц.
Всего около 40 молекул этого белка можно обнаружить в одном вирионе этого семейства вирусов. Протеин состоит из двух субъединиц, которые находятся в равном количестве и выполняют три важнейшие функции ревертазы:
1) Синтез молекулы ДНК как на матрице одноцепочечной/двухцепочечной РНК, так и на основе дезоксирибонуклеиновых кислот.
2) Активация РНКазы Н, главная роль которой заключается в расщеплении молекулы РНК в комплексе РНК-ДНК.
3) Разрушение участков молекул ДНК для встраивания в геном эукариот.
Механизм ОТ
Этапы обратной транскрипции могут варьироваться в зависимости от семейства вирусов, т.е. от вида их нуклеиновых кислот.
Рассмотрим сначала те вирусы, которые используют ревертазу. Здесь процесс ОТ делится на 3 этапа:
Такой способ размножения вирионов характерен для некоторых онкогенных вирусов и вируса иммунодефицита человека (ВИЧ).
Стоит отметить, что для синтеза любой нуклеиновой кислоты на матрице РНК нужна затравка, или праймер. Праймер представляет собой короткую последовательность нуклеотидов, комплементарную 3’-концу молекулы РНК (матрицы) и играющую важную роль в инициации синтеза.
Когда готовые двухцепочечные молекулы ДНК вирусного происхождения встраиваются в геном эукариот, запускается обычный механизм синтеза белков вириона. В итоге “захваченная” вирусом клетка становится фабрикой по производству вирионов, где в больших количествах образуются необходимые молекулы протеинов и РНК.
Повторение таких циклов приводит как к репликации генома вируса, так и к образованию мРНК, способных к синтезу белка в условиях зараженной эукариотической клетки.
Биологическое значение обратной транскрипции
Процесс ОТ имеет первостепенное значение в жизненном цикле многих вирусов (в первую очередь ретровирусов, таких как ВИЧ). РНК вириона, атаковавшего клетку эукариот, становится матрицей для синтеза первой цепочки ДНК, на которой не трудно достроить и вторую цепь.
Полученная двухцепочечная ДНК вируса встраивается в геном эукариот, что приводит к активации процессов синтеза белков вириона и появлению большого количество его копий внутри зараженной клетки. В этом и заключается основная миссия ревертазы и ОТ в целом для вируса.
У эукариот также может встречаться обратная транскрипция в контексте ретротранспозонов – мобильных генетических элементов, способных самостоятельно транспортироваться из одного участка генома в другой. Такие элементы, как считают ученые, стали причиной эволюции живых организмов.
Ретротранспозон представляет собой участок ДНК эукариот, который кодирует несколько белков. Один из них, ревертаза, принимает непосредственное участие в делокализации такого ретротраспорозона.
Использование ОТ в науке
С того момента, как удалось выделить ревертазу в чистом виде, процесс обратной транскрипции взяли на вооружение ученые-биологи. Изучение механизма ОТ и сейчас помогает прочитать последовательности важнейших белков человека.
Дело в том, что геном эукариот, в том числе и нас, содержит неинформативные участки под названием интроны. Когда с такой ДНК считывается последовательность нуклеотидов и формируется одноцепочечная РНК, последняя лишается интронов и кодирует исключительно белок. Если с помощью ревертазы на матрице РНК синтезировать ДНК, ее легко потом секвенировать и узнать порядок нуклеотидов.
Нуклеиновая кислота, которая была образована с помощью обратной транскриптазы, называется кДНК. Она часто используется в полимеразной цепной реакции (ПЦР), чтобы искусственно увеличить число копий полученной копии кДНК. Данный метод используется не только в науке, но и в медицине: лаборанты определяют сходство такой ДНК с геномами различных бактерий или вирусов из общей библиотеки. Синтез векторов и внедрение их в бактерии – одно из перспективных направлений биологии. Если с помощью ОТ формировать ДНК человека и других организмов без интронов, такие молекулы легко внедряются в геном бактерий. Так последние становятся фабриками по производству необходимых человеку веществ (например, ферментов).
Введение ………………………………………………………………………. 3
1. Значение обратной транскрипции ………………………………………… 4
2. Механизм обратной транскрипции………………………………………. 10
3. Применение обратной транскрипции в молекулярной биологии вирусов………………………………………………………………………….
13
Реферат обратная транскрипция.doc
Наконец, реовирусы содержат в качестве генома около 10 различных двухцепочечных молекул РНК, которые можно обозначить как (±)-РНК.
Принцип репродукции этих вирусов такой же, как если бы геномом явялялась двухцепочечная ДНК. Только ключевым ферментом является РНК-синтетаза.
Неоднократно транскрибируя (-)-цепи (±)-РНК, данный фермент образует (+)-цепи, выступающие в качестве мРНК. А для накопления новых двухцепочечных молекул (±)-РНК, очевидно, необходимо использование в качестве матрицы обеих цепей вирусной РНК.
Исходным материалом для ПЦР служит геномная ДНК или мРНК. В последнем случае из мРНК путем обратной транскрипции получают кДНК , которую затем используют в ПЦР. Такое исследование получило название метода ПЦР с обратной транскрипцией.
За создание метода ПЦР (полимеразная цепная реакция) Керри Мюллису в 1993 году была присуждена Нобелевская премия.
ПЦР позволяет найти в исследуемом клиническом материале небольшой участок генетической информации (несколько десятков пар нуклеотидов ДНК или РНК) любого организма, содержащийся в следовых количествах среди огромного количества нуклеотидных последовательностей иной природы, и быстро размножить его. По сути дела метод ПЦР имитирует в пробирке естественную репликацию ДНК, только повторяющуюся с огромной скоростью и столько раз, сколько это необходимо исследователю.
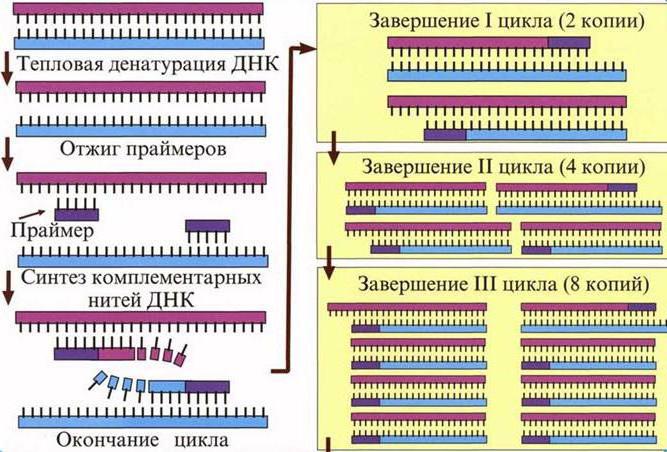
Метод включает несколько этапов: расплетание двойной спирали ДНК, расхождение нитей ДНК и последующее комплементарное дополнение (достройку) обеих с помощью специального фермента. Репликация ДНК может начаться не в любой точке, а только в определенных стартовых блоках - коротких двунитевых участках.
Для проведения такого процесса используют две генетические пробы (праймеры), которые служат в качестве затравки для синтеза второй цепи на однонитевой ДНК. Праймеры - это искусственно синтезированные короткие нуклеотидные последовательности (15-30 нуклеотидов), комплементарные концам размножаемых (амплифицируемых) участков нитей ДНК. Понятно, что, чтобы иметь нужные праймеры, необходимо знать нуклеотидную последовательность того участка ДНК, который требуется размножить. Суть метода ПЦР отображена на рисунке 5 .
Рисунок 5. Схема полимеразной цепной реакции
Сначала двунитевую ДНК нагревают до температуры около 100 град. С. При этом комплементарные нити ДНК расходятся между собой (ДНК денатурирует). Затем к обеим нитям ДНК по принципу комплементарности присоединяют искусственно синтезированные праймеры, в результате чего образуются короткие двунитевые "стартовые" участки. Далее в действие вступает специфический бактериальный фермент - Taq-полимераза, устойчивая к высоким температурам, при которых другие белки теряют свои свойства. Термоустойчивая полимераза осуществляет in vitro синтез вторых цепей ДНК на каждой из двух денатурированных цепей. После нового прогрева до 100град.С уже вновь синтезированные фрагменты ДНК служат в качестве матрицы для синтеза новых нитей в следующем цикле амплификации - это и есть цепная реакция ПЦР.
В результате такого "тиражирования" за 2-3 часа количество копий фрагмента ДНК увеличивается в геометрической прогрессии, и через 25 циклов амплификации синтезируется 10 6 копий фрагмента. Такого количества ДНК достаточно, чтобы визуально регистрировать с помощью простых приемов, которые давно используются молекулярными биологами.
Метод амплификации ДНК с помощью ПЦР (полимеразной цепной реакции) оказал революционное влияние на генодиагностику. Для использования метода необходимо знать последовательность нуклеотидов на исследуемом участке ДНК.
Поскольку ДНК-полимераза не способна сама начать репликацию и может только достраивать комплементарную цепь к уже имеющемуся двухцепочечному участку, то сначала синтезируют два олигонуклеотида (так называемых праймера), комплементарных участкам противоположных цепей ДНК, обычно отстоящих друг от друга на несколько сотен пар нуклеотидов. Праймеры инкубируют с ДНК, намеченной для амплификации, и ДНК-полимеразой, которая, как известно, синтезирует комплементарные цепи в направлении 5'-3'. Специфичность реакции зависит от правильного выбора праймеров.
В ходе реакции последовательно меняют температуру: при температуре 90-95 градусов по С происходит разделение цепей ДНК, при температуре 40-60 градусов по С - присоединение праймера (отжиг), при температуре 72 градуса по С - синтез цепей ДНК.
В реакции используют термостойкую ДНК-полимеразу, которая не теряет активности в течение всей процедуры.
После ряда таких циклов, обычно 20-30 и более, образуются сотни тысяч копий исходной последовательности, расположенной между праймерами. Метод настолько чувствителен, что его можно использовать, например, для амплификации и анализа единственного участка ДНК из одного сперматозоида человека.
Для ПЦР пригоден минимально обработанный исходный биологический материал, даже частично разрушенный, что позволяет исследовать цельную кровь, пятна высохшей крови, смывы из ротовой полости, старые срезы ткани и другие образцы. Исходным материалом для ПЦР служит геномная ДНК или мРНК. В последнем случае из мРНК путем обратной транскрипции получают кДНК , которую затем используют в ПЦР, - так называемый метод ПЦР с обратной транскрипцией .
Метод ПЦР служит основой для дальнейших исследований амплифицированной последовательности:
- гибридизации с аллель- специфическими олигонуклеотидными зондами;
- определения нуклеотидной последовательности;
- исследования экспрессии in vitro с целью поиска мутаций, укорачивающих молекулу белка.
Различные модификации метода применяются для:
- синтеза одноцепочечной ДНК путем изменения соотношения олигонуклеотидных праймеров;
- создания рекомбинантных ДНК;
- исследования мутагенеза клонированной ДНК;
- сравнения экспрессии разных аллелей;
- выявления редких нуклеотидных последовательностей или ДНК возбудителей инфекции.
ПЦР можно провести всего за один день, ее легко автоматизировать, реакция сравнительно недорога и чрезвычайно специфична.
Принцип ПЦР заключается в чередовании циклов гибридизации одноцепочечных ДНК или РНК с мечеными олигонуклеотидными зондами (праймерами), синтеза комплементарной нуклеотидной последовательности с помощью термостабильной ДНК-полимеразы и денатурации образовавшихся двухцепочечных структур путем нагревания. За 20-30 циклов количество копий нужного фрагмента нуклеиновой кислоты достигает нескольких миллионов.
Для определения продукта реакции, как правило, используют хемилюминесценцию.
§ 15. ПЕРЕНОС ГЕНЕТИЧЕСКОЙ ИНФОРМАЦИИ В КЛЕТКЕ:
РЕПЛИКАЦИЯ, ТРАНСКРИПЦИЯ
В процессе размножения клеток генетическая информация передается от одного поколения клеток другому. При этом все клетки получают одинаковую информацию. Это, возможно, вследствие того, что перед делением клетки осуществляется репликация (удвоение) ДНК, в результате образуются две идентичные молекулы ДНК, которые и передаются потомкам. В структуре ДНК заложена способность этой молекулы к копированию.
Закодированная в ДНК генетическая информация реализуется в результате экспрессии генов. Экспрессия генов включает транскрипцию (копирование информации с ДНК на синтезируемую РНК) и последующую трансляцию (синтез на матрице РНК соответствующего белка).
Возможен поток информации и в направлении от РНК к ДНК, этот процесс носит название обратная транскрипция. В то же время информация не передается от белков нуклеиновым кислотам. Однако следует отметить, что белки играют важную роль в осуществлении процессов передачи информации, как между нуклеиновыми кислотами, так и от нуклеиновых кислот к белкам.
Описанные информационные взаимоотношения между ДНК, РНК и белками могут быть представлены в виде схемы:
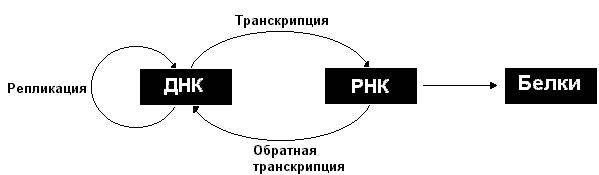
Рассмотрим более подробно процессы репликации, транскрипции и трансляции.
Интересно знать! В 1982 году Р.Д.Пальмитер с коллегами опубликовали фотографию, на которой рядом сидели две мыши. Одна из них была трансгенной, в ее ДНК встроили ген гормона роста крысы, другая была обычной мышкой. Трансгенная мышь в сравнении с обычной имела в два раза большие размеры. Причиной тому была экспрессия гена гормона роста, в результате которой у мыши синтезировался соответствующий гормон, который и определил ее гигантские размеры.
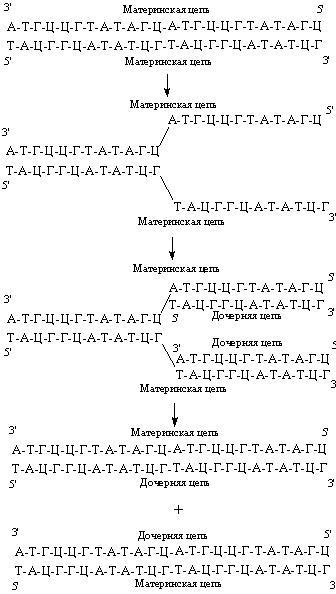
Рис. 44. В результате репликации образуются две дочерние молекулы ДНК.
Репликация ДНК
В основе репликации, или удвоения, ДНК лежит принцип комплементарности. Репликация начинается с разделения цепей, каждая из которых становится матрицей, определяющей нуклеотидную последовательность новой комплементарной цепи (рис. 44). В результате репликации образуются две дочерние молекулы ДНК, нуклеотидные последовательности которых идентичны между собой и с материнской молекулой ДНК. При этом две дочерние молекулы ДНК содержат одну вновь синтезированную (дочернюю) и одну материнскую цепи ДНК.
Субстратом для синтеза ДНК являются дезоксинуклеотид-5’-трифосфаты: дАТФ, дГТФ, дЦТФ и дТТФ. Уравнение этой реакции в простейшей виде выглядит так:
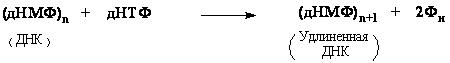
Данную реакцию катализирует фермент ДНК-полимераза. Она последовательно добавляет нуклеотиды к 3’-концу полинуклеотидной цепи. Следовательно, синтез новой цепи происходит в направлении от 5’-конца к 3’-концу. ДНК-полимераза может только наращивать цепь ДНК, начать же синтез ДНК с нуля она не может, т.е. для ее работы требуется цепь-затравка. ДНК-полимераза способна удлинять цепь только в присутствии цепи, играющей роль матрицы (рис. 45.). Нуклеотиды присоединяются к цепи-затравке в соответствии с принципом комплементарности, напротив аденина всегда будет встроен тимин, а напротив гуанина – цитозин.
Итогом репликации является образование двух дочерних молекул ДНК, являющихся точными копиями материнской. Благодаря этому возможна передача идентичной генетической информации от клетки к клетке.
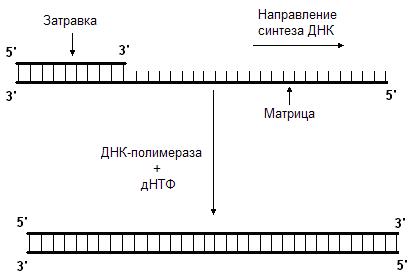
Рис. 45. ДНК-полимераза наращивает цепь в направлении 5’ 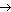
3’.
Для этого ей нужны матрица и затравка
Транскрипция
В результате транскрипции происходит синтез цепи РНК, нуклеотидная последовательность которой комплементарна одной из цепей ДНК; при этом генетическая информация, содержащаяся в ДНК, передается молекуле РНК. В результате транскрипции образуются три класса РНК: иРНК, тРНК, рРНК. Поскольку в основе транскрипции лежит принцип комплементарности, этот процесс имеет определенное сходство с репликацией. Однако между ними существует и важное различие. Если в процессе репликации копируется вся молекула ДНК, то при транскрипции транскрибируется только ее незначительная часть. Катализирует синтез РНК фермент РНК-полимераза. Транскрибируемый участок ДНК ограничен со стороны 3’-конца промотором, – участком с которым связывается РНК-полимераза, со стороны 5’-конца – терминатором – участком, в котором прерывается синтез РНК. Последовательность ДНК, ограниченная промотором и терминатором, представляет собой единицу транскрипции – транскриптон (рис. 46).
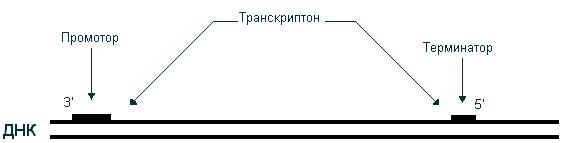
Рис. 46. Транскриптон
РНК-полимераза в качестве субстратов для синтеза РНК использует рибонуклеозид-5’-фосфаты (АТФ, ГТФ, ЦТФ и УТФ). Она удлиняет цепь, присоединяя нуклеотиды к 3’-концу, т.е. так же, как и ДНК-полимераза, наращивает цепь в направлении5’®3’. Этот фермент требует для своей работы в качестве матрицы одну из цепей ДНК. Из двух цепей ДНК транскрибируется только одна. В отличие от ДНК-полимеразы, РНК-полимераза не нуждается в затравке. Удлинение цепи РНК описывается уравнением:

Нуклеотиды присоединяются к цепи в соответствии с принципом комплементарности. Напротив аденина матричной цепи ДНК в растущую цепь РНК всегда будет встроен урацил, напротив гуанина – цитозин, напротив тимина – аденин и напротив цитозина – гуанин соответственно. Т.о., образовавшаяся в результате транскрипции РНК комплементарна матричной цепи ДНК (рис. 47).
Рис. 47. Образовавшаяся в результате транскрипции РНК комплементарна матричной цепи ДНК
В процессе транскрипции выделяют три стадии: инициацию, элонгацию и терминацию. На стадии инициации РНК-полимераза, взаимодействуя с промотором, вызывает расхождение цепей ДНК и начинает синтез молекулы РНК. В ходе элонгации новосинтезированная цепь РНК образует (за счет спаривания ее оснований с основаниями матричной цепи ДНК) короткие отрезки гибридной двойной спирали ДНК – РНК, которые необходимы для правильного считывания цепи ДНК. Как только РНК-полимераза достигнет терминирующих последовательностей, запускается последняя стадия – терминация. По ее завершении происходит освобождение вновь синтезированной цепи РНК (рис. 48).
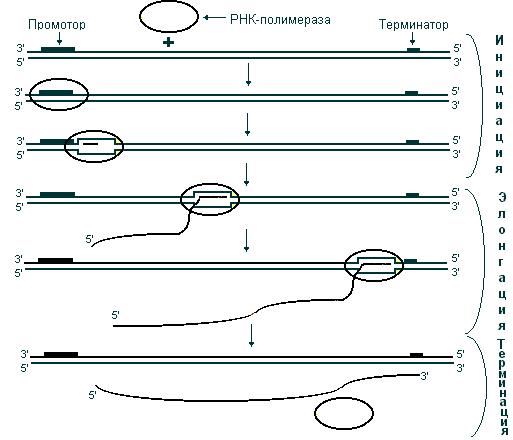
Рис. 48. Стадии транскрипции
Синтезированные молекулы РНК могут подвергаться посттранскрипционным ковалентным модификациям, так называемому процессингу (созреванию). Наиболее ярко процессинг выражен у эукариот.
Процессинг РНК
Рассмотрим процессинг иРНК эукариот.
иРНК синтезируется в виде предшественника пре-иРНК. Модификация пре-иРНК начинается на стадии элонгации, в это время происходит кэпирование 5’-конца. Остаток ГТФ присоединяется своим 5’-концом к 5’-концу пре-иРНК с образованием так называемого кэпа. После завершения транскрипции к 3’-концу присоединяется полиА-последовательность, состоящая из 100 – 200 последовательно соединенных адениновых нуклеотидов. В составе пре-иРНК могут содержаться интроны и экзоны. Интроны при созревании РНК вырезаются, концы экзонов соединяются друг с другом. Этот процесс называется сплайсингом. В результате сплайсинга зрелая молекула иРНК становится приблизительно в 4 раза короче пре-иРНК. Этапы процессинга иРНК представлены на рис. 49. Процессинг для иРНК прокариот не характерен.
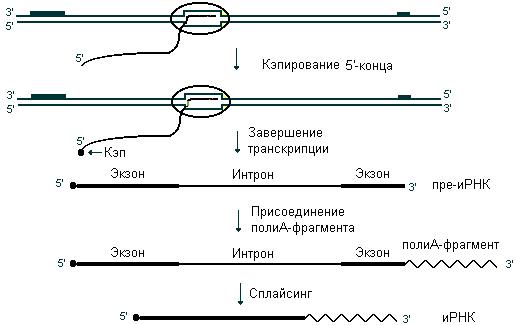
Рис. 49. Процессинг иРНК
рРНК и тРНК также синтезируются в виде более длинных предшественников, которые затем расщепляются и модифицируются. У прокариот рРНК представлены тремя молекулами: 16S-рРНК, 23S-рРНК и 5S-рРНК, у эукариот – четырьмя: 18S-рРНК, 28S-рРНК, 5,8S-рРНК и 5S-рРНК. рРНК как прокариот, так и эукариот образуются из предшественников – пре-рРНК. Предшественник расщепляется, образуя индивидуальные рРНК (рис. 50). рРНК и тРНК не содержат кэпа и полиА-последовательности. У эукариот рРНК и тРНК могут подвергаться сплайсингу. В процессе созревания у некоторых предшественников тРНК наряду с удалением концевых последовательностей может происходить и присоединение нуклеотидных последовательностей, играющих важную роль в их функционировании.
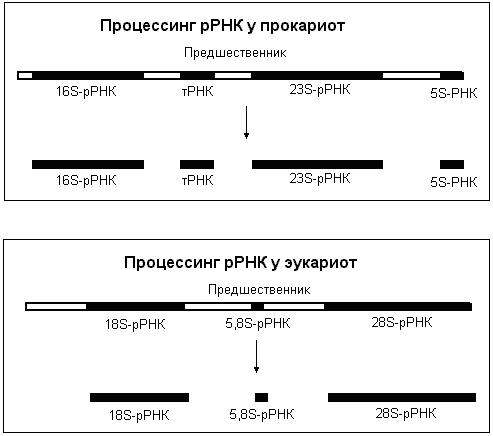
Рис.50. Процессинг рРНК.
Обратная транскрипция
Синтез ДНК при использовании в качестве матрицы РНК носит название обратной транскрипции. Данный процесс катализирует фермент обратная транскриптаза или ревертаза. Существование обратных транскриптаз в составе РНК-содержащих вирусов было показано Г.Темином и Д. Балтимором. Обнаружение обратной транскриптазы позволило ответить на вопрос: как генетическая информация РНК-содержащих вирусов может включиться в ДНК клетки-хозяина. Процесс обратной транскрипции, катализируемый ревертазой, и последующая интеграция генетического материала в геном клетки хозяина представлены на рис. 51. В процессе обратной транскрипции вначале образуется дуплекс РНК – ДНК, затем РНК в составе этого дуплекса разрушается, синтезированная цепь ДНК далее служит матрицей для синтеза второй цепи ДНК. На заключительной стадии происходит интеграция ДНК, синтезированной в результате обратной транскрипции, в ДНК клетки-хозяина. Обратные транскриптазы способны синтезировать ДНК, комплементарную самым различным РНК. Благодаря этой особенности ревертаза нашла широкое применение в научных исследованиях. С помощью обратной транскриптазы можно получить, например, искусственный ген, используя в качестве матрицы иРНК.
Читайте также:
