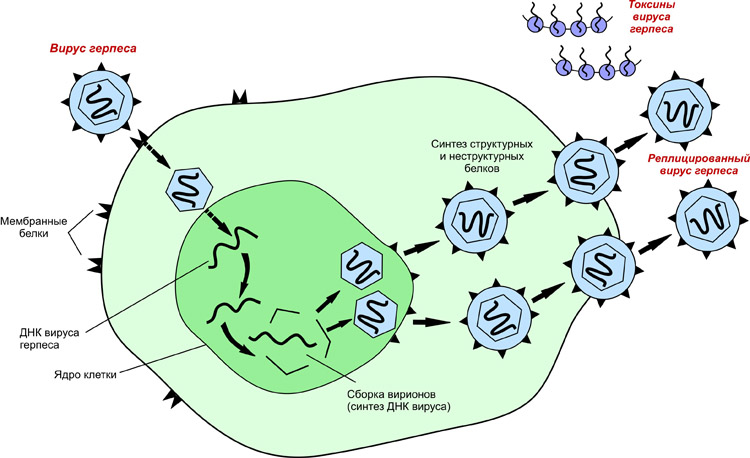
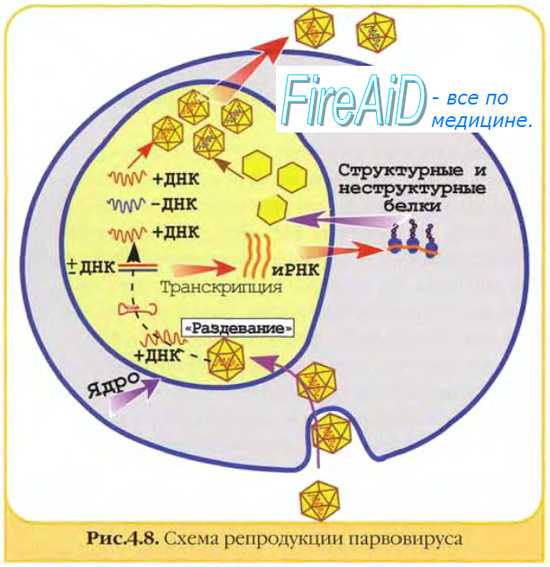
Механизмы репликации рнк-содержащих вирусов

Учебник предназначен для студентов, аспирантов и преподавателей высших медицинских учебных заведений, университетов, микробиологов всех специальностей и практических врачей.
5-е издание, исправленное и дополненное
Книга: Медицинская микробиология, иммунология и вирусология
Репликация вирусных геномов
Репликация вирусных геномов
Тип геномной вирусной ДНК определяет особенности ее репликации.
1. Двунитевая ДНК – репликация происходит по обычному механизму полуконсервативной репликации: нити разделяются, и на каждой из них достраивается комплементарная ей нить.
3. У вирусов, геном которых представлен однонитевой РНК, ее репликация происходит по следующей схеме: вначале на вирионной РНК (вРНК) синтезируются комплементарные ей РНК (кРНК). Этот процесс катализируется специфической РНК-репликазой I. Затем на кРНК синтезируется комплементарная ей, но идентичная исходной вирионная РНК (вРНК), этот процесс также катализируется специфической репликазой II. Таким образом, репликация идет по схеме (рис. 80. II):
Рис. 80. Механизм репликации однонитевого вирусного ДНК-генома (I) и однонитевого вирусного РНК-генома (II):
а – геномная однонитевая ДНК; б – репликативная форма; в – промежуточная репликативная форма; г – однонитевая дочерняя геномная ДНК; вРНК – вирионная (геномная) РНК; кРНК – комплементарная РНК. Объяснение в тексте
5. Размножение вируса гепатита В также протекает с участием обратной транскриптазы, но вначале клеточная РНК-полимераза синтезирует на вирусной ДНК прегеномную РНК, после чего вирусная ревертаза синтезирует на ней минус-цепь ДНК, которая достраивается плюс-цепью ДНК. В виде двунитевой ДНК вирус интегрируется в хромосому клетки-хозяина, где на ней транскрибируется вирусная РНК.
Существуют некоторые общие закономерности размножения вирусов. Во-первых, все РНК-содержащие вирусы, кроме вирусов гриппа и ретровирусов, размножаются в цитоплазме. Для своего размножения вирусы гриппа А и В и ретровирусы проникают в ядро, что связано с особенностями поведения их генома. Во-вторых, размножение всех ДНК-содержащих вирусов, кроме вирусов оспы, протекает в ядре, где происходит транскрипция и репликация их геномных нуклеиновых кислот, и в цитоплазме, где происходит трансляция вирусных белков, их процессинг и морфогенез вирионов. Лишь размножение вирусов группы оспы происходит в цитоплазме клетки, поскольку они обладают собственными системами транскрипции.
Другая особенность размножения вирусов заключается в том, что их нуклеокапсидные белки синтезируются на свободных полирибосомах (не связанных с мембраной), а суперкапсидные белки – на рибосомах, ассоциированных с мембранами (на шероховатых мембранах). Кроме того, белки некоторых вирусов подвергаются протеолитическому процессингу и гликозилированию. Различают два типа протеолитического процессинга: каскадный и точечный.
При точечном протеолизе разрезанию подвергается один (реже несколько) из вирусных полипептидов. Разрезание происходит, как правило, в определенном участке полипептида. Такой тип протеолиза необходим для того, чтобы определенный белок вируса приобрел свою специфическую активность. Например, суперкапсидный белок вируса гриппа – гемагглютинин – разрезается на две субъединицы: боEльшую и меньшую. В результате меньшая субъединица приобретает способность сливаться с мембранами клетки-мишени и ее лизосомами. Благодаря этому вирус гриппа приобретает способность проникать в клетку. Такой точечный протеолиз наблюдается у ортомиксовирусов, парамиксовирусов, ротавирусов, вирусов группы оспы и др. Точечный протеолиз, как и каскадный, жизненно важен для вируса.
Наконец, еще одна особенность вирусов, обладающих суперкапсидом, заключается в том, что суперкапсидные белки подвергаются в ходе своей транспортировки на наружную поверхность клеточной мембраны гликозилированию.
Информация
Добавить в ЗАКЛАДКИ| Поделиться: |
РНК вируса репликацияВероятно, вирус-сателлит сам кодирует свой собственный структурный белок, так как серологически он не родствен ВНТ. В растениях, зараженных ВНТ, не удалось обнаружить антигены, которые были бы серологически родственны ВС [1413], однако результаты опытов нельзя считать окончательными, так как свободные белковые субъединицы могут и не реагировать с антисывороткой, специфичной в отношении интактного вируса. Более веским доводом в пользу предположения, что ВС кодирует специфический структурный белок, служит отсутствие серологической связи между ВС и ВНТ, как отмечалось выше, хотя ВНТ и способствует репликации вируса-сателлита. Анализ пептидов триптического гидролизата продуктов синтеза in vitro показал, что при этом образуется материал, сходный со структурным белком ВС.[ . ] У некоторых РНК-содержащих вирусов в геноме закодированы РНК-зависимые транскриптазы (обратные транскриптазы). Поэтому репликация генома РНК-содержащих вирусов катализируется собственной обратной транскриптазой, которая упаковывается в капсиде при каждой репликации вирусов в клетке-хозяине. Такие РНК-содержащие вирусы получили название ретровирусов (от лат. retro — возврат назад).[ . ] Среди мелких РНК-содержащих вирусов животных больше всего известно о репликации вируса полиомиелита, обычно изучаемой на линии клеток человека (клетки HeLa), растущих, в суспензионной культуре. Сразу же после заражения клеток вируслая РНК высвобождается из своей оболочки. Новые вирусоспецифичдыс белки, необходимые для образования вируса, синтезируются также вскоре после заражения. В их число входит РНК-полимераза, которая, вероятно, функционально сходна с ферментом, обнаруженным у РНК-содержащих бактериофагов. Синтез большей части вирусной РНК и белка оболочки вируса происходит между 2,5 и 5 ч после заражения. В зараженной клетке образуется двухцепочечная форма вирусной РНК, и количество ее растет экспоненциально в течение первых нескольких часов после заражения. Эксперименты с мечеными предшественниками позволили установить, что синтез одной молекулы РНК вируса полиомиелита в клетках HeLa занимает около 1 мин. Вирусная РНК функционирует в качестве информационной РНК в полирибосомах, которые в зараженных клетках HeLa отличаются значительно большими размерами, чем в незараженных. По-видимому, эта вирусная PIIK функционирует как полицистронпая информационная PIIK. Вирусоспецифичные полирибосомы прикрепляются в зараженной клетке к какой-то большой, содержащей липиды структуре. Изучение вируса полиомиелита in vivo выявило, что по меньшей мере 10 различных полипептидов, пе входящих в состав оболочки, и 4 полипептида вирусной оболочки кодируются вирусной РНК [1702]. Полностью сформированные (зрелые) вирусные частицы начинают появляться приблизительно через 2,5 ч после заражения. Вслед за этим происходит быстрое (в течение 2—3 мин) включение вновь синтезированной вирусной РНК в зрелые вирусные частицы. В то же время для включения вирусного белка требуется в среднем около 20 мин. Это позволяет предполагать, что в процессе репликации вируса образуется пул вирусного белка, используемого при формировании вирусных частиц.[ . ] ДНК-содержащие вирусы имеют либо собственные ферменты репликации (в капсиде), либо в их геноме закодирована информация о синтезе вирусных ферментов, обеспечивающих репликацию вирусной нуклеиновой кислоты. Количество этих ферментов различно в применении к разным вирусам. Например, в геноме бактериального вируса Т4 закодирована информация о синтезе около 30 вирусных ферментов. Далее геном крупных вирусов кодирует нуклеазы, разрушающие ДНК клетки-хозяина, ,а также белки, воздействие которых на клеточную РНК-полимеразу сопровождается тем, что «обработанная таким образом РНК-полимераза транскрибирует на разных стадиях вирусной инфекции разные вирусные гены. Напротив, малые по размерам ДНК-содержащие вирусы в большей мере зависят от ферментов клеток-хозяев. Например, синтез ДНК аденовирусов обеспечивается клеточными ферментами.[ . ] Возбудитель — Rabdovirus carpio, РНК-содержащий, имеет пулевидную форму, размером (105—125) х (70—85) нм. Он представлен одним серотипом. Среди полевых изолятов имеет авирулентные штаммы. Вирус размножается в первично трипсинизированных культурах клеток гонад карпа и в перевиваемых линиях клеток рыб — ЕРС, FHM и др. Оптимальная температура для репликации вируса в культуре тканей 20—23 °С. Он обнаружен в крови, асцитной жидкости, почках, печени, селезенке, слизистой кишечника, мускулатуре, головном мозге.[ . ] Возбудитель — рабдовирус, РНК-содержащий, со спиральной симметрией, имеет пальцевидную форму, размер (180—240) х (60— 75) нм. Покрыт гладкой оболочкой. На его плоском конце иногда различают отросток длиной 80—90 нм. Вирус хорошо культивируется на перевиваемых линиях клеток рыб FHM и RTG-2 (культура фибропластов из яичников радужной форели), а также карповой линии ЕРС. Репликация вируса в клетках происходит при температурах 10—21 °С, достигая титра 105—106«5 ТЩЬ/мл.[ . ] Если двухцепочечная форма вирусной РНК имеет отношение к репликации вируса, то можно было бы, очевидно, ожидать, что в клетке она будет появляться раньше, чем иитактный вирус. Экспериментально, однако, это еще не было показано. Возможно, что продемонстрировать это вообще довольно трудно. С помощью заР удалось обнаружить двухцепочечную форму РНК ВЖМТ в листьях китайской капусты через 3 дня после заражения, тогда как серологические тесты не позволяли выявить присутствие вируса вплоть до 5-го дня [1379]. Этот результат почти несомненно отражает относительную чувствительность использованных методов. Применяя 32Р для обнаружения не только двухцепочечной РНК, но ВЖМТ, мы сумели выявить вирус на 2-й день после заражения, а двухцепочечную форму РНК лишь на 3-й день. Судя по содержанию РНК в клетках, можно думать, что очень небольшого количества двухцепочечной РНК достаточно для образования значительных (возможно, в 1000 раз больших) количеств вирусной РНК и эролых вирусных частиц. Задержка во времени между образованием двухцепочечной РНК и первым появлением зрелого вируса в клетке, по всей вероятности, невелика, не более нескольких минут. С другой стороны, минимальное время, необходимое для того, чтобы выявить включение 32Р в отдельные компоненты вируса, составляет 30—60 мин. Если для исследования используются ткани листа, то лшпь небольшая часть клеток оказывается инфицированной в начале периода включения 32Р; остальные клетки заражаются в значительной мере асинхронно, так что это заражение растягивается иногда на несколько дней. Таким образом, почти неизбежно первым моченым вирусным продуктом, обнаруживаемым после заражения, будет свободная одноцепочечная вирусная РНК или полный вирус, а не двухцепочечная вирусная РНК.[ . ] Бесклеточные системы с успехом используются при изучении репликации вирусов животных и бактерий. Предпринималось много попыток приспособить их также и для изучения вирусов растений. Были испробованы: 1) системы, включающие клеточные оргапеллы, окруженные мембранами; 2) неочищенные системы, не содержащие таких оргаиелл; 3) системы, содержащие частично очищенные ферменты и нуклеиновые кислоты из инфицированных растений; 4) бесклеточные системы, полученные не из растений, а из других организмов (обычно из бактерий) и содержащие в качестве информационной РНК рибонуклеиновую кислоту фитопатогенных вирусов.[ . ] Некоторое время предполагали, что явление рекомбинации ограничено только вирусами, содержащими в качестве генетического материала ДНК. Аналогичные эксперименты с вирусами растений малочисленны вследствие технических трудностей, на которые наталкиваются попытки предпринять подобного рода опыты. Тем не менее Бесту удалось получить данные, свидетельствующие о рекомбинации между штаммами вируса броязовости томатов 1185—188, 190, 191].[ . ] До самого последнего времени считалось неразумным проводить аналогии между вирусами, поражающими различные крупные группы организмов. Однако достижения молекулярной биологии продемонстрировали единство основных механизмов синтеза нуклеиновых кислот и белка в природе. Поэтому, прежде чем анализировать довольно скудцыо экспериментальные данные о размножении вирусов растений, мы кратко рассмотрим основные особенности репликации некоторых РНК-содержащих бактериофагов и вирусов животных. Даже в отношении этих вирусов детали процесса репликации изучены еще далеко не достаточно.[ . ] Анализ ВТМ и ВЖМТ из обработанных растений показал, что 8-азагуаиин включается в РНК вирусной частицы, замещая часть (несколько процентов) остатков гуанина [1174, 1176, 1177]. Вирусные препараты, содержащие аналог, были менее инфекциопны, чем нормальный вирус, и мы предположили, что ингибирующий эффект может быть обусловлен (по крайней мере частично) образованием стерильных вирусных частиц. В свете современных данных о репликации вирусов весьма маловероятпо, чтобы такой процесс лежал в основе ингибирования. Более вероятно, что аналог препятствует функционированию репликативной формы вирусной РНК или влияет па эффективность матричной функции вирусной РНК, однако пока этот вопрос не исследован.[ . ] Общий механизм, описанный выше, гораздо труднее согласовать с тем, что нам известно о вирусах, содержащих РНК. Их репликация осуществляется за счет Р Н К - з а в и с и м о г о синтеза РНК. Существование такого процесса в нормальных клетках пока не известно. Однако в препаратах ДНК-зави-симой РНК-полимеразы из Micrococcus lysodeikticus [1258] и из Azoiobacler vinelandii [1021] выявлена активность РНК-зависимой РНК-полимеразы, причем в этой системе актиномицин D не влиял на синтез РНК. Возможно, РНК-содержащие вирусы образовались из фрагментов информационной РНК клетки-хозяина, которые каким-то образом приобрели капсидный белок и способность к специфической саморепликации.[ . ] Работа с этими аналогами была начата еще до того, как появилась какая-либо возможность прошшиуть в глубь процесса репликации вируса. Трудно было представить, каким образом можно заблокировать синтез вирусной РНК, не вызвав повреждения клетки вследствие нарушения синтеза клеточных РНК. Однако теперь известно, что вирусные РНК реплицируются на РНК-матрице. Данных о том, что такой РНК-зависимый синтез РНК происходит в пезаражепной клетке, не имеется. Таким образом, по крайней мере в принципе возможно блокировать синтез вирусной РНК, не влияя на синтез клеточных РНК.[ . ] Мозаичное заболевание листьев китайской капусты, инфицированной ВЖМТ, обычно развивается при участии серии штаммов этого вируса. Мы предложили следующие гипотезы, объясняющие природу этого мозаичного заболевания [347]: 1) инфекционный вирус передвигается из ипокулирован-ного листа к зопе делящихся клеток, расположенных ниже апикальной меристемы. С помощью тестов на инфекционность было установлено, что частицы ВЖМТ присутствуют в этой зоне у системно инфицированных растений (Фаэд и Мэтьюз, неопубликованные данные); 2) вирус мутирует с образованием штаммов, оказывающих различное действие на хлоропласта клеток инфицированных растений. В полученном нами изоляте ВЖМТ мы наблюдали различные штаммы вируса, причем все попытки получить препарат, содержащий только один штамм повторным пассивированием через некроз, были безуспешны; 3) репликация первой молекулы РНК, инфицирующей данную клетку, предотвращает репродукцию других штаммов в этой клетке. При делении первой инфицированной клетки исходный штамм вируса передается дочерним клеткам, и таким образом формируется клон клеток, содержащих только один штамм вируса. Клоны образовавшихся при этом клеток дают начало островкам ткани, которые видны невооруженным глазом, а также слоям клеток, обнаруживаемым с помощью микроскопа в участках с уже развившимися симптомами мозаичного заболевания. Результаты наблюдений за развитием этого заболевания и окончательным распределением клеток, инфицированных различными штаммами вируса, свидетельствуют в пользу предположения о том, что отдельные островки ткани, наблюдаемые в инфицированных ВЖМТ листьях, представляют собой клоны клеток, происшедших от одной клетки или довольно небольшой группы клеток, зараженных каким-то одним вирусным штаммом на ранней стадии онтогенеза листа.[ . ] Подводя итоги, можно сказать, что самые различные факты, такие, как сохранение инфекционное™ после линьки, наличие латентного периода, способность к заражению в течение длительного срока и присутствие вируса в гемолимфе и клетках тела насекомого, свидетельствуют о том, что многие циркулирующие вирусы размножаются в тлях-переносчиках. В настоящее время наиболее убедительные данные получены для вирусов некротического пожелтения салата-латука, пожелтения жилок осота и скручивания листьев картофеля. Остается, однако, точно установить фат’ размножения вируса в переносчике. Для некоторых вирусов возможно получить биохимические доказательства репликации вируса в организме тли; для этого можно было бы инфицировать тлей, затем подкармливать их па здоровом растительном материале, меченном 32Р, и попытаться выделить из них меченый вирус или меченую двухцепочечпуго вирусную РНК.[ . ] Такое распределение генетического материала между несколькими компонентами должно иметь какое-то преимущество для выживания. Вероятнее всего, преимущество подобной многокомпонентной системы заключается в ее большей генетической гибкости по сравнению с вирусами, у которых весь генетический материал заключен в одном фрагменте. Известно, что у некоторых РНК-содержащих вирусов животных, в частности у реовиру-сов (гл. Многокомпонентные вирусы растений можно рассматривать как своего рода морфологический вариант такой системы, т. е. системы, в которой генетический материал разделен на несколько фрагментов. У вирусов, для которых характерно присутствие нескольких фрагментов РНК в одной частице, вероятно, происходит перераспределение генетического материала в процессе репликации. Кроме того, у таких вирусов возможны отбор и перераспределение генетического материала также па других стадиях жизненного цикла, например при передаче вируса, его проникновении в клетку и в процессе перемещения по растению.[ . ]
Вирусы — это микроскопические патогены, заражающие клетки живых организмов для самовоспроизводства. Они состоят из одного вида нуклеиновой кислоты (или ДНК или РНК, но не обе вместе), которая защищена оболочкой, содержащей белки, липиды, углеводы или их комбинацию. Размер типичного вируса варьируется от 15 до 350 нм, поэтому его можно увидеть только с помощью электронного микроскопа. В 1892 году русский ученый Д.И. Ивановский впервые доказал существование ранее неизвестного типа возбудителя болезней, это был вирус мозаичной болезни табака. А в 1898 году Фридрих Лоффлер и Пол Фрош нашли доказательства того, что причиной ящура у домашнего скота была инфекционная частица, которая меньше, чем любая бактерия. Это были первые шаги к изучению природы вирусов, генетических образований, которые лежат где-то в серой зоне между живыми и неживыми состояниями материи. На текущий момент описано около 6 тыс. вирусов, но их существует несколько миллионов. Строение вирусовВне клеток-хозяев вирусы существуют в виде белковой оболочки (капсида), иногда заключенного в белково-липидную мембрану. Капсид обволакивает собой либо ДНК, либо РНК, которая кодирует элементы вируса. Находясь в такой форме вне клетки, вирус метаболически инертен и называется вирионом. Простая структура, отсутствие органелл и собственного метаболизма позволяет некоторым вирусам кристаллизоваться, т.е. они могут вести себя подобно химическим веществам. С появлением электронных микроскопов было установлено, что их кристаллы состоят из тесно прижатых друг к другу нескольких сотен миллиардов частиц. В одном кристалле вируса полиомиелита столько частиц, что ими можно заразить не по одному разу всех жителей Земли. Формы вирусовВирусы встречаются в трех основных формах. Они бывают:
Проникновение вирусов в клетку-хозяинаКапсид в основном защищает нуклеиновую кислоту от действия клеточного нуклеазного фермента. Но некоторые белки капсида способствуют связыванию вируса с поверхностью клеток-хозяев, и работают, как ключики, вставляемые в нужные замочки. Другие поверхностные белки действуют как ферменты, они растворяют поверхностный слой клетки-хозяина и таким образом помогают проникновению нуклеиновой кислоты вируса в клетку-хозяина. Жизненный цикл вирусов сильно отличается у разных видов, но существует шесть основных этапов жизненного цикла вирусов: Присоединение к клетке-хозяину представляет собой специфическое связывание между вирусными капсидными белками и рецепторами на клеточной поверхности. Эта специфика определяет хозяина вируса. Проникновение следует за прикреплением: вирионы проникают в клетку-хозяина через рецептор-опосредованный эндоцитоз или слияние мембран. Это часто называют вирусной записью. Проникновение вирусов в клетку достигается за счет: Размножение вирусовПосле того, как вирусный геном освобождается от капсида, начинается его транскрипция или трансляция. Именно эта стадия вирусной репликации сильно различается между ДНК- и РНК-вирусами и вирусами с противоположной полярностью нуклеиновой кислоты. Этот процесс завершается синтезом новых вирусных белков и генома (точных копий внедрённых).
Механизм репликации зависит от вирусного генома.
Чтобы преобразовать РНК в ДНК, вирусы должны содержать гены, которые кодируют вирус-специфический фермент обратной транскриптазы. Она транскрибирует матрицу РНК в ДНК. Обратная транскрипция никогда не происходит в неинфицированных клетках. Необходимый фермент, обратная транскриптаза, происходит только от экспрессии вирусных генов в инфицированных клетках. ВироидыВироиды заражают только растения. Одни вызывают экономически важные заболевания сельскохозяйственных культур, в то время как другие являются доброкачественными. Двумя примерами экономически важных вироидов являются кокосный cadang-cadang (он вызывает массовую гибель кокосовых пальм) и вироид рубцовой кожицы яблок, который безнадежно портит товарный вид яблок. 30 известных вироидов были классифицированы в две семьи.
В отличие от вирусов, которые являются паразитами механизма трансляции хозяина, вироиды являются паразитами клеточных транскрипционных белков. Бактериофаги
Существуют тысячи разновидностей фагов, каждый из которых может заразить только один тип или несколько близких типов бактерий или архей. Фаги классифицируются по ряду семейств вирусов; например: Как и все вирусы, фаги являются простыми организмами, которые состоят из ядра генетического материала (нуклеиновой кислоты), окруженного капсидом белка. Нуклеиновая кислота может представлять собой либо ДНК, либо РНК, и может быть двухцепочечной или одноцепочечной. Существует три основных структурных формы фага:
Во время заражения фаг прикрепляется к бактерии и вставляет в нее свой генетический материал. После этого фаг обычно следует одному из двух жизненных циклов: литическому (вирулентному) или лизогенному (умеренному). Литические, или вирулентные, фаги захватывают механизм клетки, чтобы скопировать компоненты фага. Затем они разрушают или лизируют клетку, высвобождая новые частицы фага. Лизогенные, или умеренные, фаги включают свою нуклеиновую кислоту в хромосому клетки-хозяина и реплицируются с ней как единое целое, не разрушая клетку. При определенных условиях лизогенные фаги могут индуцироваться в соответствии с литическим циклом. Существуют и другие жизненные циклы, в т.ч. псевдолизогенез и хроническая инфекция. При псевдолизогении бактериофаг проникает в клетку, но не использует механизм репликации клеток и не интегрируется в геном хозяина, просто как бы прячется внутри бактерии, не нанося ей никакого вреда. Псевдолизогенез возникает, когда клетка-хозяин сталкивается с неблагоприятными условиями роста и, по-видимому, играет важную роль в выживании фага, обеспечивая сохранение генома фага до тех пор, пока условия роста хозяина снова не станут благоприятными. При хронической инфекции новые фаговые частицы образуются непрерывно и длительно, но без явного уничтожения клеток. Вскоре после открытия фаги начали использовать для лечения бактериальных заболеваний человека, таких как бубонная чума и холера. Но фаговая терапия тогда не была успешной, и после открытия антибиотиков в 1940-х годах она была практически заброшена. Однако с появлением устойчивых к антибиотикам бактерий терапевтическому потенциалу фагов уделяется все больше внимания. Наше время с антибиотиками заканчивается. В 2016 году женщина в штате Невада умерла от бактериальной инфекции, вызванной Klebsiella pneumoniae, которая была устойчивой ко всем известным антибиотикам. Бактерии, устойчивые к колистину, антибиотику последней инстанции, были обнаружены на свинофермах в Китае. В настоящее время бактерии приспосабливаются к антибиотикам быстрее, чем когда-либо. Покажите ножницы которыми вирусы разрезают молекулу РНК что бы встроиться для мутации.Может что нибудь придумаете другое.К примеру деление цепочка аминокислот получив энергию из вне как одноименные заряды распадается на две. К каждой соединятся только те какие были ранее (другие проскочат мимо),казалось бы копии,но внутренняя энергия разная(уменьшается увеличивается) поэтому распад и создание. Вся химия углерода на этом построена 1000 орган соединений создает у других хим элементов этого свойства нет. Иммунная система делает накладку(интерференция)с помощью энергии интерферонов пытаясь разрушить цепочку РНК вируса.Надо помочь организму но не вакциной(вирус быстро мутирует) Вирусы, лишённые суперкапсида (например, аденовирусы) проникают в клетки путём виропексиса, а имеющие таковой (покс- и герпесвирусы) — за счёт слияния суперкапсида с клеточной мембраной. Репродуктивный цикл ДНК-содержащих вирусов включает раннюю и позднюю стадии (рис. 5-4). У крупных ДНК-вирусов имеется явное несоответствие между кодирующе ёмкостью генома и молекулярной массой вирусиндуцированных белков и белков, входящих состав вирионов. Например, у герпесвирусов лишь 15% ДНК кодирует все белки вирионов и их предшественников. Возможно, значительная часть генома содержит гены, кодирующие синтез ферментов и регуляторных белков. Папова-, адено- и герпесвирусы репродуцируются относительно однотипно, в то время как репродукция поксвирусов имеет некоторые особенности. Регуляторные белки выполняют различные функции. При заражении клетки они блокируют синтез клеточных РНК, ДНК и белка и одновременно способствуют экспрессии вирусного генома, изменяя специфичность реагирования клеточных полимераз и полирибосом. Они так же запускают репликацию клеточной ДНК, модифицированной встроенными геномами ДНК содержащих вирусов и ретровирусов, то есть репликацию вирусных геномов. Вирусспецифические полимеразы. В репликацию вирусных геномов также вовлечены вирусоспецифические ДНК-полимеразы, участвующие в образовании молекул ДНК дочерних популяций.
Матричные белкинеобходимы для репликации нуклеиновых кислот и сборки дочерних популяций. Они образуют электронно-плотные скопления в клетке, известные как тельца включений (например, тельца Гварнери при натуральной оспе). Паповавирусы и аденовирусы. Репродукция паповавирусов. Репродукция аденовирусов. Адсорбция, проникновение и депротеинизация аналогичны таковым у РНК-содержащих вирусов, но у папова- и аденовирусов депротеинизация протекает в ядре, а у РНК-вирусов — в цитоплазме.
начальный период включает стадии адсорбции вируса на клетке, проникновения в клетку, дезинтеграции (депротеинизации) или "раздевания" вируса. Вирусная нуклеиновая кислота была доставлена в соответствующие клеточные структуры и под действием лизосомальных ферментов клетки освобождается от защитных белковых оболочек. В итоге формируется уникальная биологическая структура: инфицированная клетка содержит 2 генома (собственный и вирусный) и 1 синтетический аппарат (клеточный); • после этого начинается вторая группа процессов репродукции вируса, включающая средний и заключительный периоды, во время которых происходят репрессия клеточного и экспрессия вирусного генома. Репрессию клеточного генома обеспечивают низкомолекулярные регуляторные белки типа гистонов, синтезируемые в любой клетке. При вирусной инфекции этот процесс усиливается, теперь клетка представляет собой структуру, в которой генетический аппарат представлен вирусным геномом, а синтетический аппарат — синтетическими системами клетки. 2. Дальнейшее течение событий в клетке направлено на репликацию вирусной нуклеиновой кислоты(синтез генетического материала для новых вирионов) и реализацию содержащейся в ней генетической информации (синтез белковых компонентов для новых вирионов). У ДНК-содержащих вирусов, как в прокариотиче-ских, так и в эукариотических клетках, репликация вирусной ДНК происходит при участии клеточной ДНК-зависимой ДНК-полимеразы. При этом у однонитевых ДНК-содержащих вирусов сначала образуется комплементарная нить — так называемая репликативная форма, которая служит матрицей для дочерних молекул ДНК. 3. Реализация генетической информации вируса, содержащейся в ДНК, происходит следующим образом: при участии ДНК-зависимой РНК-полимеразы синтезируются и-РНК, которые поступают на рибосомы клетки, где и синтезируются вирусспе-цифические белки. У двунитевых ДНК-содержащих вирусов, геном которых транскрибируется в цитоплазме клетки хозяина, это собственный геномный белок. Вирусы, геномы которых транскрибируются в ядре клетки, используют содержащуюся там клеточную ДНК-зависимую РНК-полимеразу. У РНК-содержащих вирусов процессы репликацииих генома, транскрипции и трансляции генетической информации осуществляются иными путями. Репликация вирусных РНК, как минус-, так и плюс-нитей, осуществляется через репликативную форму РНК (комплементарную исходной), синтез которой обеспечивает РНК-зависимая РНК-полимераза — это геномный белок, который есть у всех РНК-содержащих вирусов. Репликативная форма РНК минус-нитевых вирусов (плюс-нить) служит не только матрицей для синтеза дочерних молекул вирусной РНК (минус-нитей), но и выполняет функции и-РНК, т. е. идет на рибосомы и обеспечивает синтез вирусных белков (трансляцию). У плюс-нитевыхРНК-содержащих вирусов функцию трансляции выполняют ее копии, синтез которых осуществляется через репликативную форму (минус-нить) при участии вирусных РНК-зависимых РНК-полимераз. У некоторых РНК-содержащих вирусов (реовирусы) имеется совершенно уникальный механизм транскрипции. Он обеспечивается специфическим вирусным ферментом — ревертазой (обратной транскриптазой) и называется обратной транскрипцией. Суть ее состоит в том, что вначале на матрице вирусной РНК при участии обратной транскрипции образуется транскрипт, представляющий собой одну нить ДНК. На нем с помощью клеточной ДНК-зависимой ДНК-полимеразы синтезируется ,вторая нить и формируется двунитевой ДНК-транскрипт. С него обычным путем через образование и-РНК происходит реализация информации вирусного генома. Результатом описанных процессов репликации, транскрипции и трансляции является образование дочерних молекул вирусной нуклеиновой кислоты и вирусных белков, закодированных в геноме вируса. После этого наступает третий, заключительный период взаимодействия вируса и клетки. Из структурных компонентов (нуклеиновых кислот и белков) на мембранах цитоплазматического ретикулума клетки собираются новые вирионы. Клетка, геном которой был репрессирован (подавлен), обычно гибнет. Вновь сформировавшиеся вирионы пассивно (в результате гибели клетки) или активно(путем почкования) покидают клетку и оказываются в окружающей ее среде. Таким образом, синтез вирусных нуклеиновых кислот и белков и сборка новых вирионов происходят в определенной последовательности (разобщены во времени) и в разных структурах клетки (разобщен в пространстве), в связи с чем способ репродукции вирусов и был назван дизъюнктивным (разобщенным). При абортивной вирусной инфекции процесс взаимодействия вируса с клеткой по тем или иным причинам прерывается до того, как произошло подавление клеточного генома. Очевидно, что в этом случае генетическая информация вируса реализована не будет и репродукции вируса не происходит, а клетка сохраняет свои функции неизменными. При латентной вирусной инфекции в клетке одновременно функционируют оба генома, а при вирус-индуцированных трансформациях вирусный геном становится частью клеточного, функционирует и наследуется вместе с ним. Дата добавления: 2018-05-09 ; просмотров: 467 ; Читайте также:
 Пожалуйста, не занимайтесь самолечением!При симпотмах заболевания - обратитесь к врачу. Пожалуйста, не занимайтесь самолечением!При симпотмах заболевания - обратитесь к врачу.
|