
Литический цикл развития вирусов
Лизогенный цикл, также называемый лизогенией, является стадией процесса размножения некоторых вирусов, в основном тех, которые поражают бактерии. В этом цикле вирус вставляет свою нуклеиновую кислоту в геном бактерии-хозяина..
Этот цикл образует вместе с литическим циклом два основных механизма репликации вируса. Когда бактериофаг во время лизогенного цикла вставляет свою ДНК в бактериальный геном, он становится профаном.
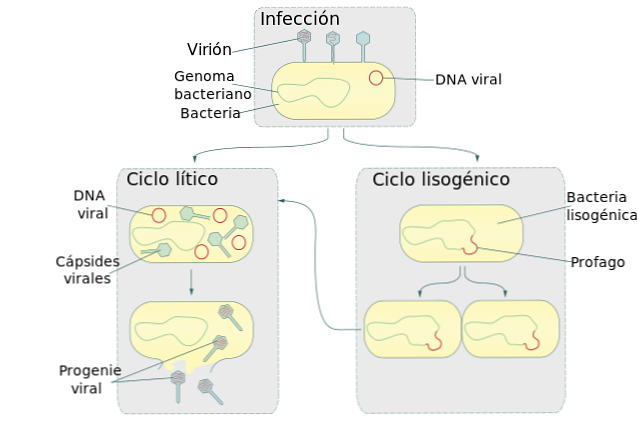
Бактерия, зараженная этим профаном, продолжает жить и размножаться. Когда происходит размножение бактерий, также получается копия профага. В результате каждая бактериальная дочерняя клетка также заражается профаном.
Размножение зараженных бактерий и, следовательно, их профагов-хозяев может продолжаться в течение нескольких поколений без проявления вируса..
Иногда самопроизвольно или в условиях стресса окружающей среды ДНК вируса отделяется от бактериальной. Когда происходит разделение бактериального генома, вирус запускает литический цикл.
Эта репродуктивная стадия вируса вызовет разрыв бактериальной клетки (лизис), что приведет к выпуску новых копий вируса. Эукариотические клетки также подвержены атаке лизогенных вирусов. Однако пока неизвестно, как происходит вставка вирусной ДНК в геном эукариотической клетки..
- 1 бактериофаг
- 2 Цикл вирусной инфекции
- 2.1 Литический цикл
- 2.2 Лизогенный цикл
- 2.3 Непрерывный цикл разработки
- 2.4 Псевдолизеногенный цикл
- 3 Лизогенная конверсия
- 4 Фаготерапия
- 4.1 Преимущества фаготерапии
- 5 ссылок
Бактериофаг
Вирусы, которые заражают только бактерии, называются бактериофагами. Они также известны как фаги. Размер этого типа вируса довольно изменчив, с диапазоном размеров, который может быть приблизительно между 20 и 200 нм..
Бактериофаги распространены повсеместно и способны развиваться практически в любой среде, где обнаружены бактерии. Подсчитано, например, что чуть менее трех четвертей бактерий, населяющих море, заражены фагами.
Цикл вирусной инфекции
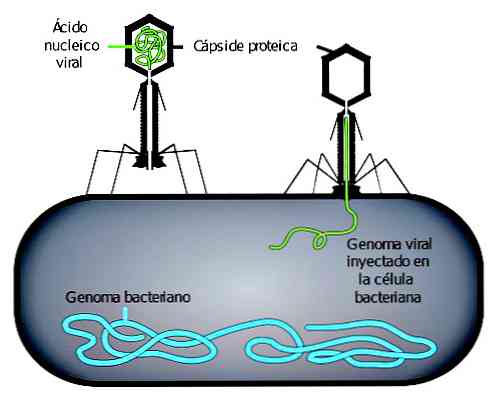
Вирусная инфекция начинается с адсорбции фага. Фаговая адсорбция происходит в два этапа. В первом, известном как обратимое, взаимодействие между вирусом и его потенциальным хозяином слабое.
Любое изменение условий окружающей среды может привести к прекращению этого взаимодействия. Вместо этого в необратимом взаимодействии участвуют специфические рецепторы, которые предотвращают прерывание взаимодействия..
ДНК вируса может проникать внутрь бактерий только тогда, когда происходит необратимое взаимодействие. Впоследствии, в зависимости от типа фага, они могут выполнять различные репродуктивные циклы.
В дополнение к уже описанным литическим и лизогенным циклам существуют два других репродуктивных цикла: непрерывное развитие и псевдолиогенный..
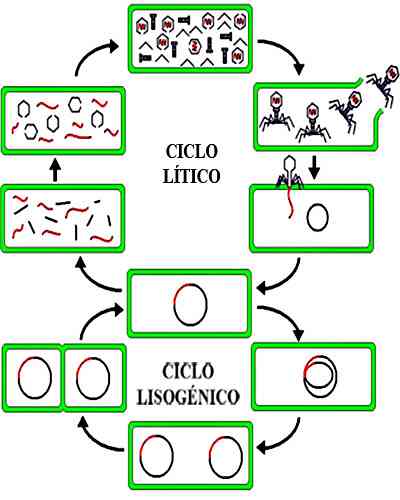
На этом этапе размножение вируса внутри бактерий происходит быстро. В конце концов бактерии будут страдать от лизиса клеточной стенки и новые вирусы будут выпущены в окружающую среду..
Каждый из этих недавно выпущенных фагов может атаковать новую бактерию. Последовательное повторение этого процесса позволяет экспоненциально расти инфекции. Бактериофаги, которые участвуют в литическом цикле, называются вирулентными фагами..
В этом цикле лизис клетки-хозяина не происходит, как это происходит в литическом цикле. После стадий адсорбции и проникновения стадия интеграции фаговой ДНК в бактериальную клетку продолжается и превращается в профаго..
Репликация фага будет происходить одновременно с размножением бактерий. Profagos, интегрированный в бактериальный геном, будет наследоваться дочерними бактериями. Вирус может продолжаться без проявления в течение нескольких поколений бактерий..
Этот процесс является частым, когда количество бактериофагов велико по сравнению с количеством бактерий. Вирусы, которые осуществляют лизогенный цикл, не являются вирулентными и называются умеренными..
В конце концов, profagos можно отделить от бактериального генома и трансформировать в литические фаги. Последние входят в литогенный цикл, что приводит к бактериальному лизису и заражению новых бактерий.
Некоторые бактериофаги выполняют многочисленные репликации внутри бактерий. В этом случае, вопреки тому, что происходит во время лизогенного цикла, он не вызывает бактериального лизиса..
Вновь реплицированные вирусы высвобождаются из бактерий в определенных местах на клеточной мембране, не вызывая их разрушения. Этот цикл называется непрерывным развитием.
Иногда доступность питательных веществ в окружающей среде недостаточна для нормального роста и размножения бактерий. В этих случаях считается, что доступной клеточной энергии недостаточно, чтобы фаги вызывали лизогению или лизис..
Из-за этого вирусы попадают в псевдолизеногенный цикл. Этот цикл, однако, еще мало известен.
Лизогенная конверсия
В конце концов, продукт взаимодействия между профаго и бактерией, первый может вызвать появление изменений в фенотипе бактерии..
Это происходит главным образом, когда бактерия-хозяин не является частью обычного цикла вируса. Это явление называется лизогенной конверсией.
Изменения, вызванные в бактериях ДНК профага, увеличивают биологический успех хозяина. Увеличивая биологическую способность и успех выживания бактерий, вирус также приносит пользу..
Этот тип выгодных отношений для обоих участников может быть классифицирован как тип симбиоза. Однако мы должны помнить, что вирусы не считаются живыми существами..
Основным преимуществом лизогенно-трансформированных бактерий является их защита от нападения других бактериофагов. Лизогенная конверсия может также увеличить патогенность бактерий у их хозяев..
Даже непатогенные бактерии могут стать патогенными вследствие лизогенного превращения. Это изменение в геноме является постоянным и наследуемым.
лечение обильной диета
Фаготерапия - это терапия, которая включает применение фага в качестве контрольного механизма для предотвращения распространения патогенных бактерий. Эта методология бактериального контроля была впервые использована в 1919 году..
В этом случае она использовалась для лечения пациента, страдающего дизентерией, и получила совершенно благоприятный результат. Фаготерапия была успешно использована в начале прошлого века.
С открытием пенициллина, а также других антибиотиков субстанция фаготерапии практически прекратилась в Западной Европе и на американском континенте..
Беспорядочное использование антибиотиков позволило создать штаммы бактерий, мультирезистентные к антибиотикам. Эти бактерии становятся все более частыми и устойчивыми.
В связи с этим в западном мире появился новый интерес к развитию фаготерапии для контроля за контаминацией и бактериальными инфекциями..
1) Рост фага происходит экспоненциально, усиливая его действие со временем, антибиотики наоборот, со временем теряют свое действие из-за метаболического разрушения молекулы.
2) Фаги обладают способностью подвергаться мутациям, что позволяет им бороться с сопротивлением, которое бактерии могут развить к их атаке. Напротив, антибиотики всегда имеют один и тот же активный ингредиент, поэтому, когда бактерии развивают устойчивость к таким активным ингредиентам, антибиотики бесполезны
3) Фаготерапия не имеет побочных эффектов, которые могут быть вредны для пациентов..
4) Разработка нового штамма фага является гораздо более быстрой и дешевой процедурой, чем открытие и разработка нового антибиотика..
5) Антибиотики воздействуют не только на патогенные бактерии, но и на другие потенциально полезные. Фаги, с другой стороны, могут быть видоспецифичными, так что лечение против бактерий, ответственных за инфекцию, может быть ограничено, не затрагивая другие микроорганизмы..
6) Антибиотики не убивают все бактерии, поэтому выжившие бактерии могут передавать генетическую информацию, которая придает устойчивость к антибиотику их потомкам, создавая таким образом устойчивые штаммы. Лизогенетические бактериофаги убивают бактерии, которые они заражают, уменьшая возможность развития устойчивых штаммов бактерий..
Общая характеристика вирусов.
Вирусы - уникальные агенты на нашей планете. Основные свойства вирусов, по которым они отличаются от всех остальных живых существ следующие:
1. Ультрамикроскопические размеры
2. Вирусы содержат нуклеиновую кислоту только одного вида - или ДНК, или РНК. Все другие организмы содержат нуклеиновые кислоты обоих типов, а геном у них представлен только ДНК.
3. Вирусы не способны к росту и бинарному делению.
4. Вирусы размножаются путем воспроизводства себя из собственной геномной нуклеиновой кислоты. Все прочие организмы способны к увеличению своей биомассы (росту) и размножаются путем бинарного деления клеток.
5. У вирусов отсутствуют собственные системы мобилизации энергии.
6. У вирусов нет собственных белоксинтезирующих систем.
7. В связи с отсутствием собственных систем синтеза белка и мобилизации энергии вирусы являются абсолютными внутриклеточными паразитами. Средой обитания вирусов являются бактерии, клетки растений, животных и человека.
С учетом перечисленных особенностей можно дать следующее определение:
Вирусы - особое царство ультрамикроскопических размеров организмов, обладающих только одним типом нуклеиновых кислот, лишенных собственных систем синтеза белка и мобилизации энергии и являющихся поэтому абсолютными внутриклеточными паразитами.
Молекулярно-генетическая организация вирусов.
Основой таксономии вирусов является вирион, который представляет собой конечную фазу развития вируса. Размеры вирионов различных вирусов варьируют в широких пределах: от 15 -18 до 300-400 нм. Они имеют разнообразную форму - палочковидную, нитевидную, сферическую. Вирион состоит из из геномной нуклеиновой кислоты, окруженной одной или двумя оболочками.
Оболочка, в которую упакована геномная нуклеиновая кислота, называется капсидом (от греческого сapsa - ящик). Наиболее просто организованные вирусы представляют собой нуклеокапсиды: они состоят только из нуклеиновой кислоты и белковой оболочки, построенной из идентичных пептидных молекул. Капсид имеет строго упорядоченную структуру, в основе которой лежат принципы спиральной или кубической симметрии.
Капсиды палочковидных или нитевидных вирионов состоят из структурных субъединиц, уложенных в виде спирали вокруг оси. При таком расположении субъединиц образуется полый канал, внутри которого компактно уложена молекула вирусной нуклеиновой кислоты. Ее длина может во много раз превышать длину палочковидного вириона. Например, длина вируса табачной мозаики 300 нм, а его РНК достигает величины 4000 нм. При этом РНК настолько связана с капсидом, что ее нельзя при этом освободить, не повредив последний. Подобные капсиды встречаются у некоторых бактериальных вирусов и у вирусов человека (вирус гриппа).
Сферическая структура вирионов определяется капсидом, построенном по принципам кубической симметрии в основе которой лежит фигура икосаэдра -двадцатигранника .
Капсид состоит из асимметричных субъединиц (полипептидных молекул), которые объединены в морфологические субъединицы - капсомеры.Один капсомер содержит 2,3,5 субъединиц, расположенных по осям симметрии икосаэдра.
Число капсомеров для вируса данного вида является постоянным, оно имеет диагностическое значение. Например, вирион аденовирусов имеет 252 капсомера, у паповавирусов - 72. Молекулярная организация всех простых вирусов сводится к использованию спиральной и кубической симметрии.
Более сложно устроены вирусы, у которых имеется вторая оболочка. Она называется - суперкапсид. Суперкапсид представляет собой обычную биологическую мембрану, состоящую из двух слоев липидов, имеющих клеточное происхождение и заключенных в них гликозилированных суперкапсидных вирусных белков, которые выступают над наружной поверхностью вириона в виде своеобразных шипов. Суперкапсидные вирусные белки, образующие шипы, обладают жизненно важными для вируса функциями:
они распознают клеточные рецепторы и связываются с ними, обеспечивают слияние вирусной мембраны с мембраной клетки и ее лизисом, способствуют распространению вируса в организме за счет слияния клеток, многие из них обладают свойствами протективных антигенов и т.д. Многие сложные вирусы, такие как ортомиксовирусы, парамиксовирусы, коронавирусы и др. устроены таким образом, что их нуклеокапсид, имеющий палочковидную спиральную структуру, окружен суперкапсидной липопротеидной оболочкой, придающей вириону сферическую форму. У тогавирусов нуклеокапсид имеет форму икосаэдра, который окружен суперкапсидной оболочкой, придающей вириону шаровидную форму. Вирион ретровирусов имеет икосаэдрический капсид внутри которого располагается спиральный нуклеокапсид, а
сам вирион покрыт липидсодержащей оболочкой, придающей ему сферическую форму.
Классификация и таксономия вирусов.
Вирусы составляют царство Vira , которое подразделяется на два подцарства по типу нуклеиновой кислоты: ДНК-содержащие вирусы (дезоксирибовирусы) и РНК- содержащие вирусы ( рибовирусы).
Подцарства делятся на семейства, которые в свою очередь подразделяются на роды. Название всех вирусных родов оканчивается словом "virus", для названия семейств используется суффикс "idae", а подсемейств "inae". Из более чем 55 семейств вирусов, признанных Международным комитетом по таксономии вирусов, 19 включают вирусы человека и животных.
Для классификации вирусов используют следующие критерии:
1. Нуклеиновая кислота: тип, число нитей, процентное содержание, молекулярная масса, содержание гуанина и цитозина.
2. Морфология: тип симметрии или псевдосимметрии, число капсомеров для вирусов с кубической симметрией, наличие внешней липопротеиновой оболочки, форма. Размеры вирионов.
3. Биофизические свойства.
4. Белки: количество структурных белков, их локализация , аминокислотный состав.
6. Размножение в тканевых культурах.
7. Круг поражаемых хозяев, особенности патогенеза инфекционного процесса
8. Устойчивость к физическим и химическим факторам (гамма-лучи, термоинактивация при 37 и 50 градусах Цельсия, действие жирорастворителей и отдельных катионов).
9. Антигенные свойства.
В состав простых вирионов входит один тип нуклеиновой кислоты - РНК или ДНК и белки. У сложных вирионов в составе внешней оболочки содержатся липиды и полисахариды, которые они получают из клеток хозяина.
Вирусные ДНК. По своей структуре вирусные ДНК характеризуются рядом особенностей, что дает возможность подразделить их на несколько типов:
1. Одноцепочечная линейная ДНК (парвовирусы)
2. Одноцепочечная кольцевая ДНК
3. Двухцепочечная линейная ДНК (вирусы герпеса)
4. Двухцепочечная кольцевая ДНК (паповавирусы, вирус гепатита В)
5. Двухцепочечная ДНК с ковалентно связанным терминальным гидрофобным белком ( аденовирусы)
6. Двухцепочечная ДНК, замкнутая на каждом конце ковалентной связью (вирус оспы).
Вирусные РНК. У РНК-содержащих вирусов генетическая информация закодирована в РНК таким же кодом. Как в ДНК всех других вирусов и клеточных организмов. Вирусные РНК по своему химическому составу не отличаются от РНК клеточного происхождения, на характеризуются разной структурой:
1. Одноцепочечная нефрагментированная РНК, обладающая матричной активностью (позитивная или +РНК) Вирус полиомиелита и др. пикорнавирусы.
2. Одноцепочечная нефрагментированная РНК, не обладающая матричной активностью (негативная, или - РНК). Вирион имеет в своем составе РНК-зависимую РНК-полимеразу (транскриптазу). Она синтезирует на вирионной РНК матричную РНК, необходимую для трансляции вирусспецифических белков. Парамиксовирусы.
3. Одноцепочечная фрагментированная РНК, не обладающая матричной активностью (негативная РНК), вирион имеет транскриптазу. Ортомиксовирусы (РНК состоит из 8 фрагментов).
4. Двухцепочечная фрагментированная РНК, вирион имеет транскриптазу. Реовирусы.
5. Вирусы, геном которых представлен двумя идентичными нитями позитивной РНК (диплоидный геном). Вирионы имеют обратную транскриптазу. Ретровирусы.
6. Одноцепочечная кольцевая РНК. Такой геном имеет только один вирус - вирус Дельта - гепатита. Это дефектный вирус, для его размножения необходим вирус - помощник - вирус гепатита В.
Жизненный цикл вирусов. Основные типы вирусных геномов.
Под жизненым циклом вируса понимают процесс его размножения. Он происходит только внутриклеточно. Особенности размножения зависят прежде всего от вирусного генома.
Тип вирусной ДНК определяет особенности ее репликации.
1. Двунитевая ДНК - репликация происходит по обычному механизму репликации: нити разделяются, и на каждой из них достраивается комплиментарная ей нить.
2. Однонитевая ДНК. Ее репликация происходит через образование вначале репликативной формы, а затем промежуточной репликативной формы. Репликативная форма возникает в результате синтеза на исходной вирионной ДНК ( + нити) комплементарной ей (-) нити, то есть однонитевая ДНК превращается в двунитевую структуру ДНК. Промежуточная репликативная форма - это репликативная форма, (-) нить которой служит матрицей для синтеза (+) нити ДНК, идентичной исходной вирионной ДНК. Такой механизм обеспечивает передачу генов дочерним вирионам.
3. У вирусов, геном которых представлен однонитевой РНК, ее репликация происходит по следующей схеме: вначале на вирионной РНК (в РНК) синтезируются комплиментарные ей РНК (к РНК). Этот процесс катализирется специфической РНК - репликазой 1. Затем на к РНК синтезируется комплементарная ей, но идентичная исходной вирионная РНК (в РНК), этот процесс также катализируется специфической репликазой П. Таким образом репликация идет по схеме :
в РНК - - к РНК -- в РНК
4. Репликация однонитевой РНК ретровирусов происходит с участием обратной транскриптазы. Вначале на в РНК обратная транскриптаза синтезирует комплементарную ей "минус" цепь ДНК, а затем на ней - "плюс" нить ДНК. Двунитевая ДНК интегрируется в хромосому клетки и там служит матрицей для синтеза разных классов вирусных РНК. Таким образом, репликация ретровирусов происходит по схеме:
Существуют некоторые общие закономерности размножения вирусов. Во- первых, все РНК-содержащие вирусы, кроме вирусов гриппа и ретровирусов, размножаются в цитоплазме.
Во-вторых, размножение всех ДНК-содержащих вирусов, кроме вирусов оспы, протекает в ядре.
Другая особенность размножения вирусов заключается в том, что их нуклекапсидные белки синтезируются на свободных полирибосомах, (не связанных с мембраной), а суперкапсидные белки - на рибосомах, ассоциированных с мембранами. Кроме того, белки некоторых вирусов подвергаются протеолитическому процессингу и гликозилированию.
Вирусы - это наименьшие структуры, обладающие свойствами живого организма. Их размер составляет от 20 до 300 нм в длину. Они невидимы в оптический микроскоп и легко проходят через фильтры, задерживающие бактерии.
Согласно классификации организмов вирусы находятся на границе живого и неживого. Их считают живыми, так как они содержат генетический материал и способны размножаться; в то же время их можно считать неживыми, поскольку они не имеют клеточной организации - вирусы неклеточные формы жизни. Вне клетки хозяина вирусы не проявляют признаков жизни.
Вирусы и болезни. Поскольку вирусы являются облигатными паразитами, они неизбежно наносят вред клетке-хозяину, вызывая ряд серьезных заболеваний. Вирусные заболевания часто с трудом поддаются лечению, поскольку антибиотики в данном случае не действуют (у вирусов нет метаболических процессов, которые можно было бы ингибировать); вакцины не всегда эффективны, так как вирусы мутируют, изменяя свои антигенные свойства; химиотерапия может не только ингибировать репликацию вируса, но и принести вред самим клеткам. Обычно используются такие формы контроля, как вакцинация и удаление источника заражения. Некоторые ретровирусы могут играть роль в лечении заболеваний - они могут быть использованы для переноса в хозяйскую клетку ДНК, содержащей дефектную форму определенного гена (таким путем может быть излечена, например, ФКУ - фенилкетонурия).
Обычный вирус представляет собой простую структуру, состоящую из кора (сердцевины), содержащего нуклеиновую кислоту (ДНК или РНК), и белковой оболочки, окружающей сердцевину.

это область, ограниченная капсидом. Он не является цитоплазмой и не содержит органелл.
Выросты на поверхности
капсида отвечают за антигенные свойства вируса
вируса может быть представлен либо ДНК, либо РНК. Количество генов очень невелико - в них содержится только информация, необходимая для репликации вирусных субъединиц и для сборки из них целой вирусной частицы, или вириона.
состоит из множества идентичных субъединиц, называемых капсомерами. Они самоорганизуются в строго симметричный капсид, форма которого используется для классификации вирусов.
окружает капсид у некоторых вирусов, обычно у крупных. Эта оболочка чаще всего образуется из элементов клетки-хозяина, формируясь в процессе выхода вириона из клетки. Она может играть важную роль в обеспечении способности вируса преодолевать защитные барьеры клетки. Такую оболочку имеет, например, вирус иммунодефицита человека (ВИЧ).

Вирусы используют ресурсы клетки-хозяина для образования многочисленных копий самих себя, и их сборка происходит внутри клетки. процесс делится на несколько этапов:
Этапы жизненного цикла вируса
Образование связи между белками вирусного капсида и рецепторами на поверхности клетки-хозяина. Эта связь определяет круг хозяев вируса, то есть инфицирование вирусом только тех клеток, которые способны осуществить его репликацию. иИзменения белка оболочки служит сигналом к проникновению вируса в клетку.
Проникновение в клетку
Вирус доставляет внутрь клетки свой генетический материал (иногда собственные белки). Разные вирусы используют разные стратегии.
Процесс потери каспида при помощи вирусных ферментов или клетки-хозяина, либо результат обычной диссоциации.
Репликация вируса - включает синтез мРНК ранних генов вируса. Синтез вирусных белков, сборка сложных белков и репликацию вирусного генома.
Сборка вирусных частиц, затем модификация белков.
Выход из клетки
Вирусы могут покинуть клетку после лизиса, процесса, в ходе которого клетка погибает из-за разрыва мембраны и клеточной стенки. Активно размножающийся вирус не всегда убивает клетку-хозяина. Оболочечные вирусы (например ВИЧ) обычно отделяются от клетки путём отпочковывания.
_______________
Источник информации: Биология человека в диаграммах / В.Р. Пикеринг — 2003.






Строение и жизненный цикл вирусов
Вирусы представляют собой неклеточную форму жизни. Они неспособны к самостоятельному размножению и обмену веществ, поэтому для реализаций этих функций вирусам необходима клетка-хозяин. Вирусы были обнаружены двадцативосьмилетним русским ученым Д.И. Ивановским в 1892 г . Еще будучи студентом Петербургского университета (1887), Д.И. Ивановский начал изучать причины, вызывающие заболевание табака, при котором на листьях последнего появлялась мозаика.
Строение вирусов. Как мы уже говорили, вирусы не имеют клеточного строения. Каждая вирусная частица устроена очень просто - она состоит из расположенного в центре носителя генетической информации и оболочки. Генетический материал представляет собой короткую молекулу нуклеиновой кислоты, это образует сердцевину вируса. Нуклеиновая кислота у разных вирусов может быть представлена ДНК или РНК, причем эти молекулы могут иметь необычное строение: встречается однонитчатая ДНК и двухнитчатая РНК. Оболочка называется капсид. Она образована субъединицами - капсомерами, каждый из которых состоит из одной или двух белковых молекул. Число капсомеров для каждого вируса строго постоянно (например, в капсиде вируса полиомиелита их 60 - не больше и не меньше, а у вируса табачной мозаики - 2130, причем не 2129 и не 2131). Иногда нуклеиновая кислота вместе с капсидом называется нуклеокапсидом. Если вирусная частица, кроме капсида, больше не имеет оболочки, ее называют простым вирусом, если имеется еще одна - наружная, вирус называется сложным. Наружную оболочку также называют суперкапсидом, генетически она не принадлежит вирусу, а происходит из плазматической мембраны клетки-хозяина и формируется при выходе собранной вирусной частицы из инфицированной клетки. Таким образом, вирусная частица состоит только из двух классов биополимеров: нуклеиновых кислот и белков, тогда как в любой клетке в обязательном порядке должны присутствовать еще полисахариды и липиды.
У каждого вируса капсомеры капсида располагаются в строго определенном порядке, благодаря чему возникает определенный тип симметрии. При спиральной симметрии капсид приобретает трубчатую (вирус табачной мозаики) или сферическую (РНК-содержащие вирусы животных) форму. При кубической симметрии капсид имеет форму икосаэдра (двадцатигранника), такой симметрией обладают изометрические вирусы. В случае комбинированной симметрии капсид обладает кубической формой, а расположенная внутри нуклеиновая кислота уложена спирально. Правильная геометрия капсида даже позволяет вирусным частицам совместно образовывать кристаллические структуры.
Жизненный цикл вирусов. Вирусы не могут самостоятельно раз-множаться и осуществлять обмен веществ. В соответствии с этим у них различают две жизненные формы: покоящаяся внеклеточная -вирион и активно репродуцирующаяся внутриклеточная - вегетативная. Вирионы демонстрируют отменную жизнеспособность. В частности, они выдерживают давление до 6000 атм и переносят высокие дозы радиации, однако погибают при высокой температуре, облучении ультрафиолетовыми лучами, а также воздействии кислот и дезинфицирующих веществ. Взаимоотношения вируса с клеткой последовательно проходят несколько стадий (рис. 1).
![]()
Первая стадия представляет собой адсорбцию вирионов на поверхности клетки-мишени, которая для этого должна обладать соответствующими поверхностными рецепторами. Именно с ними специфически взаимодействует вирусная частица, после чего происходит их прочное связывание, по этой причине клетки восприимчивы не ко всем вирусам. Именно этим объясняется строгая определенность путей проникновения вирусов. Например, рецепторы к вирусу гриппа имеются у клеток слизистой оболочки верхних дыхательных путей, а у клеток кожи их нет. Поэтому через кожу гриппом заболеть нельзя - вирусные частицы для этого нужно вдохнуть с воздухом. Вирусы бактерий (бактериофаги) нитевидной формы или не имеющие отростков адсорбируются не на клеточной стенке, а на фимбриях. Вначале вирионы адсорбируются посредством электростатического взаимодействия или за счет ван-дер-ваальсовых сил (собственно поэтому вирусы осаждаются не только на поверхности клеток, но и на любой поверхности вообще). Первая фаза адсорбции обратима - вирусную частицу можно отделить от клетки-мишени (например, обычным встряхиванием), после чего следует необратимая фаза, при которой разделение уже невозможно.


Третья стадия называется депротеинизация. В ходе ее происходит освобождение носителя генетической информации вируса - его нуклеиновой кислоты. У многих вирусов, например бактериофагов (за исключением нитчатых), этот процесс совпадает с предыдущей стадией, поскольку в клетку проникает только нуклеиновая кислота, а белковая оболочка остается за пределами клетки-хозяина. Если вирус проникает в клетку целиком, то удаление оболочки осуществляется клеточными протеазами. Напомним, что вирион может проникать в клетку в результате эндоцитоза. Как и положено, при этом формируется вакуоль-фагосома, с которой сливаются первичные лизосомы. Однако в случае обычного фаго- или пиноцитоза ферменты лизосом расщепляют органические вещества фагосомы до мономеров, которые впоследствии используются клеткой для своих нужд. По невыясненным до конца причинам с проникшими в клетку вирионами этого не происходит в полной мере. Ферментативному расщеплению подвергается лишь белковая составляющая вирусной частицы, а его нуклеиновая кислота остается неповрежденной. В результате нуклеиновая кислота вируса освобождается, и впоследствии именно она существенным образом преобразует деятельность клетки-хозяина, подчиняя ее метаболизм своим потребностям и вынуждая ее синтезировать определенные вещества. Обращаем внимание на то, что сам вирус не обладает необходимыми для этого механизмами, поэтому для синтеза нужных молекул он использует клеточные ферменты (например, протеазы, РНК-полимеразы и др.) и структуры (например, рибосомы). Пути реализации генетической информации разными вирусами называют стратегией вирусного генома.
РНК-содержащие вирусы по этому признаку делятся на несколько групп. Наиболее просто все устроено у представителей первой группы (пикорна-, тога- и коронавирусы). У них транскрипция не происходит, потому что вирионная однонитчатая РНК сама выполняет функцию мРНК, т.е. служит матрицей для синтеза вирионного белка на рибосомах клетки. Следовательно, схема биосинтеза у них следующая: РНК -> белок. Такие вирусы называются плюснитевые (или вирусы с позитивным геномом).
Вторую группу составляют минус-нитевые вирусы (или вирусы с негативным геномом - вирусы гриппа, кори, паротита и др.). Они также содержат однонитчатую РНК, однако она не информативна для прямой трансляции, поэтому у них сначала происходит транскрипция на вирионной РНК комплементарной ей m-РНК, которая и будет служить матрицей для последующего синтеза вирус-ных белков. Следует отметить, что транскрипция управляется ферментом РНК-зависимой РНК-полимеразой. Этот фермент отсутствует в клетке (понятно, что клетке он просто не нужен, поскольку в ней никогда не синтезируется РНК на РНК) и приносится самим вирионом. В этом случае схема биосинтеза будет: РНК -> РНК -> белок.
У составляющих третью группу ретровирусов (они относятся к онковирусам) биосинтез идет наиболее сложно. У них на исходной однонитчатой РНК-матрице сначала синтезируется ДНК - уникальный случай в природе, которому нет аналогов. Этот процесс управляется особым ферментом - РНК-зависимой ДНК-полимеразой (его еще называют обратной транскриптазой, или ревертазой). Полученная молекула ДНК впоследствии приобретает кольцевую форму и называется ДНК-провирус. Затем эта молекула встраивается в хромосому клетки-хозяина и с помощью клеточной же РНК-полимеразы многократно транскрибируется. Образовавшиеся копии выполняют следующие функции: во-первых, являются мРНК, по которой на клеточных рибосомах синтезируются белки капсида вируса, во-вторых, они сами непосредственно являются РНК вириона. Таким образом, схема биосинтеза у этих вирусов: РНК -> ДНК -> РНК -> белок.
Четвертую группу образуют вирусы, содержащие двухнитчатую РНК. У них транскрипция также осуществляется с помощью вирусного фермента РНК-зависимой РНК-полимеразы.
В пятой стадии происходит синтез компонентов вирусной частицы - нуклеиновой кислоты и белков капсида, причем все компоненты синтезируются многократно.
В ходе шестой стадии из синтезированных ранее многочисленных копий нуклеиновой кислоты и белков формируются новые вирионы путем самосборки. Для этого необходимо, чтобы концентрация компонентов вириона достигла высокого (критического) уровня. Обращаем внимание на то обстоятельство, что компоненты вирусной частицы синтезируются раздельно и в разных частях клетки. У сложных вирусов, кроме капсида, также образуется наружная оболочка из компонентов плазматической мембраны клетки.
Последняя - седьмая стадия - представляет собой выход вновь собранных вирусных частиц из клетки-хозяина. У разных вирусов этот процесс проходит неодинаково. У некоторых вирусов это сопровождается гибелью клетки за счет освобождения литических ферментов лизосом - лизис клетки. У других вирионы выходят из живой клетки путем отпочковывания (см. рис. 1), однако и в этом случае клетка со временем погибнет, поскольку при отпочковывании повреждается плазматическая мембрана.
Время, прошедшее с момента проникновения вируса в клетку до выхода новых вирионов, называется скрытым, или латентным, периодом. Оно может широко варьировать: от нескольких часов (пяти-шести у вирусов оспы и гриппа) до нескольких суток (вирусы кори, аденовирусы и др.).
У некоторых бактериофагов наряду с вирулентными (быстро развивающиеся вирусы) имеются умеренные фаги. Их нуклеиновая кислота после проникновения в бактериальную клетку интегрируется в ДНК клетки и становится профагом. Профаг не оказывает литического воздействия на клетку-хозяина и при делении реплицируется вместе с клеточной ДНК. Бактерии, содержащие профаг, называются лизогенными. Они проявляют устойчивость к содержащемуся в них фагу, а также к близким к нему другим фагам. Связь профага с бактерией весьма прочная, но она может быть нарушена под воздействием индуцирующих факторов (ультрафиолетовые лучи и ионизирующая радиация, химические мутагены). Следует отметить, что лизогенные бактерии могут менять свойства (например, выделять новые токсины).
Источник: Г.Л. Билич,В.А. Крыжановский "Биология для поступающих в вузы"
Читайте также:
