
Хромосомы вирусов и прокариот
Наследственность и изменчивость в живой природе существуют благодаря генам и хромосомам, дезоксирибонуклеиновой кислоте (ДНК). Хранится и передается генетическая информация в виде цепочки нуклеотидов в составе ДНК. Какая роль в этом явлении принадлежит генам? Что такое хромосома с точки зрения передачи наследственных признаков? Ответы на подобные вопросы позволяют разобраться в принципах кодирования и генетическом разнообразии на нашей планете. Во многом оно зависит от того, сколько хромосом входит в набор, от рекомбинации этих структур.
Американский исследователь Уолтер Саттон выяснил, сколько хромосом содержится в клеточном ядре. Ученый считал эти структуры носителями единиц наследственности, признаков организма. Саттон обнаружил, что хромосомы состоят из генов, с помощью которых потомкам от родителей передаются свойства и функции. Генетик в своих публикациях дал описания хромосомных пар, их движения в процессе деления клеточного ядра. Независимо от американского коллеги, работы в том же направлении вел Теодор Бовери. Оба исследователя в своих трудах изучали вопросы передачи наследственных признаков, сформулировали основные положения о роли хромосом (1902-1903). Дальнейшая разработка теории Бовери-Саттона происходила в лаборатории нобелевского лауреата Томаса Моргана. Выдающийся американский биолог и его помощники установили ряд закономерностей размещения генов в хромосоме, разработали цитологическую базу, объясняющую механизм законов Грегора Менделя — отца-основателя генетики.
Хромосомы — наиважнейший элемент клетки. Они отвечают за передачу и реализацию наследственной информации и в эукариотической клетке локализуются в ядре. По химическому строению хромосомы представляют собой комплексы дезоксирибонуклеиновых кислот (ДНК) и связанных с ними белков, а также небольшого количества других веществ и ионов. Таким образом, хромосомы являются дезоксирибонуклеопротеидами (ДНП).
Каждая хромосома в интерфазе включает одну длинную двухцепочечную молекулу ДНК. Ген — это последовательность определенного количество следующих друг за другом нуклеотид, составляющих ДНК. Гены, входящие в состав ДНК одной хромосомы, следуют друг за другом. В интерфазе в клетке протекает множество процессов, многие участки хромосомы деспирализованы в разной степени. На многих участках ДНК идет синтез РНК.
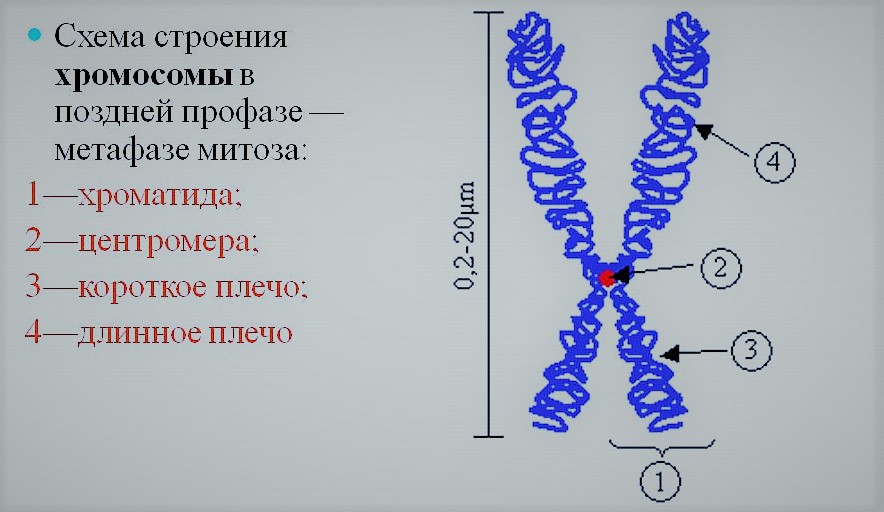
Строение удвоенной хромосомы. 1 — хроматида; 2 — центромера; 3 — короткое плечо; 4 — длинное плечо.
В период клеточного деления (как при митозе, так и при мейозе) хромосомы спирализуются (происходит их компактизация). При этом их длина сокращается, а синтез на них РНК становится невозможным. До спирализации каждая хромосома удваивается. Говорят, что хромосома становится состоящей из двух хроматид. То есть в период интерфазы хромосома состояла из одной хроматиды.
В компактизации хроматид важную роль играют белки, входящие в состав хромосомы.
Таким образом, в зависимости от фазы клеточного цикла по внешнему строению хромосомы могут быть представлены
- в виде невидимого в световой микроскоп хроматина (в интерфазе) и состоять из одной хроматиды
- в форме двух спирализованных хроматид, видимых в световой микроскоп (в фазах клеточного деления, начиная с метафазы).
В строении хромосом есть еще один важный элемент — центромера (первичная перетяжка). Она имеет белковую природу и отвечает за движение хромосомы, также к ней крепятся нити веретена деления. В зависимости от места расположения центромеры различают равноплечие (метацентрические), неравноплечие (субметацентрические) и палочковидные (акроцентрические) хромосомы. У первых центромера находится по-середине, разделяя каждую хроматиду на два равных плеча, у вторых плечи неравной длины, а у третьих центромера находится у одного из концов хроматиды.
В удвоенных хромосомах хроматиды соединены между собой в области центромеры. Наличие первичной перетяжки в строении хромосом обязательно. Однако кроме них бывают вторичные перетяжки (ядрышковые организаторы), они наблюдаются не у всех хромосом. В ядре на вторичных перетяжках хромосом происходит синтез ядрышек. На концах хроматид находятся так называемые теломеры. Они препятствуют слипанию хромосом.
В гаплоидном наборе каждая хромосома по своему строению уникальна. Положение центромеры (и обусловленные этим длины плеч хромосомы) позволяет отличать каждую среди остальных. В диплоидном наборе у каждой хромосомы есть гомологичная ей, имеющая такое же строение и тот же набор генов (но возможно других их аллелей) и доставшаяся от другого родителя.
Для каждого вида живых организмов характерен свой кариотип, т. е. свое количество хромосом и их особенности (длина, положение центромер, особенности химического строения). По кариотипу можно определить биологический вид.
Каждая хромосома обладает индивидуальным строением, отличается от других особенностями окрашивания. При изучении морфологии важно определить положение центромеры, длину и размещение плеч относительно перетяжки. В набор хромосом обычно входят следующие формы: метацентрические, или равноплечие, для которых характерно срединное расположение центромеры; субметацентрические, или неравноплечие (перетяжка смещена в сторону одного из теломеров); акроцентрические, или палочковидные, в них центромера находится практически на конце хромосомы; точковые с трудно поддающейся определению формой. Функции хромосом Хромосомы состоят из генов — функциональных единиц наследственности. Теломеры — концы плеч хромосомы. Эти специализированные элементы служат для защиты от повреждения, препятствуют слипанию фрагментов. Центромера выполняет свои задачи при удвоении хромосом. На ней есть кинетохор, именно к нему крепятся структуры веретена деления. Каждая пара хромосом индивидуальна по месту расположения центромеры. Нити веретена деления работают таким образом, что в дочерние клетки отходит по одной хромосоме, а не обе. Равномерное удвоение в процессе деления обеспечивают точки начала репликации. Дупликация каждой хромосомы начинается одновременно в нескольких таких точках, что заметно ускоряет весь процесс деления.
Выяснить, что такое хромосома, какую функцию выполняет эта ядерная структура, удалось после изучения ее биохимического состава и свойств. В эукариотических клетках ядерные хромосомы образованы конденсированным веществом — хроматином. По данным анализа, в его состав входят высокомолекулярные органические вещества: дезоксирибонуклеиновая кислота (ДНК); рибонуклеиновая кислота (РНК); белки-гистоны.
Нуклеиновые кислоты принимают самое непосредственное участие в биосинтезе аминокислот и белков, обеспечивают передачу наследственных признаков из поколения в поколение. ДНК содержится в ядре эукариотической клетки, РНК сосредоточена в цитоплазме. Гены Рентгеноструктурный анализ показал, что ДНК образует двойную спираль, цепи которой состоят из нуклеотидов. Они представляют собой углевод дезоксирибозу, фосфатную группу и одно из четырех азотистых оснований: А — аденин. Г — гуанин. Т — тимин. Ц — цитозин. Участки спиралевидных дезоксирибонуклеопротеидных нитей — это гены, несущие закодированную информацию о последовательности аминокислот в белках или РНК. При размножении наследственные признаки от родителей потомству передаются в виде аллелей генов. Они определяют функционирование, рост и развитие конкретного организма. По мнению ряда исследователей, те участки ДНК, что не кодируют полипептиды, выполняют регулирующие функции. Геном человека может насчитывать до 30 тыс. генов. Набор хромосом Общее число хромосом, их особенности — характерный признак вида. У мухи-дрозофилы их количество — 8, у приматов — 48, у человека — 46. Это число является постоянным для клеток организмов, которые относятся к одному виду.
Генетические черты связаны с половой принадлежностью. Для их передачи служат Y и Х-хромосома у мужчин, две X у женщин. Аутосомы содержат оставшуюся часть информации о наследственных признаках. Существуют методики, позволяющие индивидуализировать все 23 пары. Они хорошо различимы на рисунках, когда окрашены в определенный цвет. Заметно, что 22-я хромосома в геноме человека — самая маленькая. Ее ДНК в растянутом состоянии имеет длину 1,5 см и насчитывает 48 млн пар азотистых оснований. Специальные белки гистоны из состава хроматина выполняют сжатие, после чего нить занимает в тысячи раз меньше места в ядре клетки. Под электронным микроскопом гистоны в интерфазном ядре напоминают бусы, нанизанные на нить ДНК.
Существует более 3 тыс. наследственных болезней разного типа, обусловленных повреждениями и нарушениями в хромосомах. К их числу относится синдром Дауна. Для ребенка с таким генетическим заболеванием характерно отставание в умственном и физическом развитии. При муковисцидозе происходит сбой в функциях желез внешней секреции. Нарушение ведет к проблемам с потоотделением, выделению и накоплению слизи в организме. Она затрудняет работу легких, может привести к удушью и летальному исходу. Нарушение цветового зрения — дальтонизм — невосприимчивость к некоторым частям цветового спектра. Гемофилия приводит к ослаблению свертываемости крови. Непереносимость лактозы не позволяет организму человека усваивать молочный сахар. В кабинетах планирования семьи можно узнать о предрасположенности к тому или иному генетическому заболеванию. В крупных медицинских центрах есть возможность пройти соответствующее обследование и лечение.
Генотерапия — направление современной медицины, выяснение генетической причины наследственных заболеваний и ее устранение. С помощью новейших методов в патологические клетки вместо нарушенных вводят нормальные гены. В таком случае врачи избавляют больного не от симптомов, а от причин, вызвавших заболевание. Проводится только коррекция соматических клеток, методы генной терапии пока не применяются массово по отношению к половым клеткам.
Вирусы – неклеточные формы жизни.
Вирусы являются облигатными паразитами — они не способны размножаться вне клетки. В настоящее время известны вирусы, размножающиеся в клетках растений, животных, грибов и бактерий (последних обычно называют бактериофагами). Обнаружен также вирус, поражающий другие вирусы.
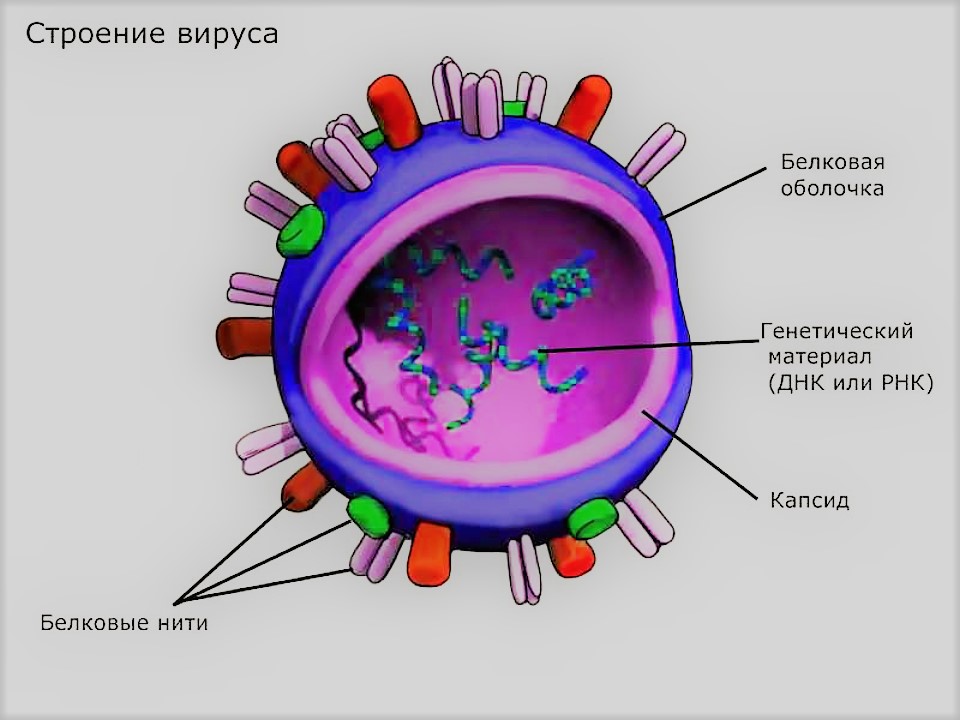
Вирусы представляют собой молекулы нуклеиновых кислот (ДНК или РНК), заключённые в защитную белковую оболочку (капсид). Наличие капсида отличает вирусы от других инфекционных агентов, вироидов. Вирусы содержат только один тип нуклеиновой кислоты: либо ДНК, либо РНК. Ранее к вирусам также ошибочно относили прионы, однако впоследствии оказалось, что эти возбудители представляют собой особые белки и не содержат нуклеиновых кислот.
Размер 0,015 – 0,350 мкм. Открыты (вирусы табачной мозаики) Д. И. Ивановским в 1892 г. Вирус активен только в клетке хозяина, в нем клетке он не имеет признаков живого организма.
Вирусные частицы (вирионы) представляют собой белковую капсулу — капсид, содержащую геном вируса, представленный одной или несколькими молекулами ДНК или РНК. Капсид построен из капсомеров — белковых комплексов, состоящих, в свою очередь, из протомеров. Нуклеиновая кислота в комплексе с белками обозначается термином нуклеокапсид. Некоторые вирусы имеют также внешнюю липидную оболочку. Размеры различных вирусов колеблются от 20 (пикорнавирусы) до 500 (мимивирусы) и более нанометров. Вирионы часто имеют правильную геометрическую форму (икосаэдр, цилиндр).
- РНК-вирусы: одноцепочечные и двуцепочечные РНК-вирусы (большинство вирусов) — СПИД, грипп, корь и т.д.
- ДНК-вирусы: одноцепочечные и двуцепочечные ДНК-вирусы — натуральная оспа
- Вирусы растений: происходит разрушение хлоропласта, обесцвечивание листьев, неспособность к фотосинтезу — мозаичная болезнь табака, гороха
- Вирусы человека и животных – свинка, бешенство, полиомиелит
- Вирусы бактерий и микроорганизмов – бактериофаг
- Генная инженерия
- Нанотехнологии
- Биологическое оружие
Одна из серьезных проблем, с которой сталкиваются разработки в области нанотехнологии, — это разброс размеров компонентов. Действительно, получить идентичные нанообъекты чрезвычайно сложно. Нанотрубки и нанопроволочки, предлагаемые в качестве элементов наноэлектронных приборов и устройств, несмотря на все усилия, имеют отличия по форме и/или размеру. Недавно был предложен новый подход к решению этой проблемы. Он заключается в использовании биологических объектов, имеющих заложенные природой строго определенные размеры и форму. К успеху привели вирусы, прекрасно работающие в качестве наноматриц для производства органических и неорганических наноматериалов или устройств.
Генетический материал эукариот имеет сложную надмолекулярную организацию, определяющую формирование хромосом. Кроме того, в клетках различных организмов обнаружены экстрахромосомные (внеядерные, цитоплазматические) молекулы ДНК.
Хромосомы вирусов и прокариот
Генетический материал зрелой вирусной частицы (вириона) представлен одной молекулой нуклеиновой кислоты (ДНК либо РНК), которая окружена защитной белковой оболочкой (капсидом). Наряду с вирусами, содержащими двухцепочечную либо одноцепочечную молекулу ДНК в замкнутой (кольцевой) форме, имеются представители, у которых эта молекула является незамкнутой (линейной) структурой и также может быть двухцепочечной либо одноцепочечной. В случае РНК-содсржащих вирусов известны как одноцепочечные, так и двухцепочечные варианты молекул РНК. Размеры хромосом таких вирусов значительно меньше, чем у большинства ДНК-co держащих вирусов, и чаше всего варьируют в пределах от 3 000 до 7 000 рибонуклеотидов.
Молекулы вирусных ДНК как правило имеют длину от 0,5 до 100 микрометров (мкм), тогда как длина вирионов обычно колеблется от 0,02 до 0,3 мкм. Столь явное несоответствие между размерами генетического материала и вмещающего его белкового капсида устраняется путем многократного закручивания молекулы ДНК вокруг оси ее спирали, что приводит к образованию значительно более короткой по длине суперспирализованной (суперзакрученной) структуры, формирующей хромосому вируса (рис. 5.32).

Рис. 5.32. Супсрспирализованная молекула ДНК: а - принцип строения суперзакрученной молекулы ДНК, превращающейся в открытую кольцевую форму при разрыве одной цепочки молекулы; б - электронная микрофотография открытой и закрытых молекул ДНК бактериофага
В результате образования супервитков линейные размеры вирусной хромосомы оказываются в несколько сотен раз меньше, чем размеры составляющей ее ДНК. Это наглядно демонстрирует электронно-микроскопическое изображение бактериофага (фага) Т2 после его последовательной обработки гипертоническим солевым раствором и дистиллированной водой (осмотического шока) (рис. 5.33). Такая обработка привела к разрыву белкового капсида фага и освобождению его молекулы ДНК, которая ранее была плотно упакована в хромосомную структуру. Измеренная длина этой молекулы составила около 50 мкм, что более чем в 500 раз превышает размеры фаговой головки, в которой она находилась.

Рис. 5.33. Электронная микрофотография молекулы ДНК бактериофага Т2, подвергнутого действию осмотического шока
Весьма незначительные размеры геномов мелких вирусов позволяют им кодировать лишь единичные белки, синтезируемые метаболической системой клетки-хозяина, в которую проник генетический материал вируса.
Так, например, в РНК самого мелкого из известных вирусов, выделяемого при некрозе листьев табака, которая состоит из 1 200 рибонуклеотидов, был обнаружен лишь один структурный ген, кодирующий белок оболочки этого вируса. Вместе с тем ДНК мелкого фага 3 пар нуклеотидов) благодаря наличию в ней перекрывающихся генов кодирует 9 различных белков, обеспечивающих формирование зрелых фаговых частиц. С другой стороны, хромосомная ДНК достаточно крупного фага Т4 (1,8 х 10 5 пар нуклеотидов) могла бы содержать не менее 100 структурных генов среднего размера (идентифицировано более 40 белков этого фага, синтезируемых зараженными клетками Е. со И).
Вирусы обладают специфичностью в отношении клеток организмов- хозяев (бактерий, растений, животных, человека), в которых они могут размножаться. После проникновения генетического материала вируса в клетку в ней начинается процесс синтеза вирусных белков и нуклеиновой кислоты на основе генетической программы этого вируса и с помощью метаболической системы хозяина. При этом проникшая РНК ретровирусов служит матрицей для синтеза комплементарной вирусной ДНК по механизму обратной транскрипции, тогда как у других РНК-содержащих вирусов может происходить лишь копирование их первичного генетического материала (молекул РНК).
Возможны два основных варианта развития вируса в клетке-хозяине, которые в случае бактериальных фагов принято называть литическим н лизогенным циклами (рис. 5.34). Развитие вирулентных вирусов (фагов) обычно происходит только по литическому пути, связанному с синтезом компонентов вирусных частиц (белков, нуклеиновой кислоты), сборкой ви- рионов и последующим разрушением (лизисом) клетки-хозяина. Однако для умеренных (латентных) вирусов, примером которых является бактериофаг X, возможен и второй вариант развития, состоящий в лизогенизации клетки-хозяина (рис. 5.34). При этом ДНК вируса интегрируется в хромосому хозяина по механизму сайт-специфической рекомбинации и существует в ней в форме провируса (профага), реплицируясь и передаваясь в процессе клеточного деления дочерним клеткам как составная часть хромосомы материнской клетки. Вместе с тем под влиянием ряда индуцирующих факторов (ультрафиолет, ионизирующая радиация, различные химические агенты и др.) происходит выход вируса из интегрированного состояния, т. е. из формы провируса (профага), в автономное состояние, что приводит к его размножению и лизису клетки-хозяина (рис. 5.34). Иными словами, возможен переход с пути лизогенизации на литический путь развития вируса и обратно. Аналогичные пути развития известны и в случае ряда вирусов эукариотических клеток, в том числе для ретровируса иммунодефицита человека, являющегося причиной возникновения СПИДа.

Рис. 5.34. Два варианта возможного развития умеренного фага X в клетках Е. coli
В отличие от вирусов хромосома бактериальной клетки содержит молекулу ДНК лишь одного типа, а именно кольцевую двухцепочечную ДНК гораздо более значительных размеров (1 000-2 000 мкм длиной). Поскольку у бактерий нет настоящего ядра, то их генетический материал организован в виде ядерноподобной структуры (нуклеоида), располагающейся в цитоплазме клетки. Каждому нуклеоиду соответствует одна хромосома, т. е. бактерии являются гаплоидными организмами.
Как и у вирусов, линейные размеры генетического материала бактерий явно не соответствуют размерам структурного образования, в котором он находится. Так, например, типичные клетки Е. coli имеют форму палочек длиной 1-5 мкм и толщиной 0,4-0,8 мкм. Это несоответствие также устраняется путем супере пир ализации молекулы ДНК, формирующей хромосому нуклеоида.
В процессе упаковки молекулы участвуют несколько ДНК-связывающих белков, обнаруженных в клетках Е. coli, часть которых по своему аминокислотному составу напоминают гистоны из хромосомы эукариот.
Кроме того, в нуклеоидах этих бактерий найдена РНК, роль которой остается неясной. Посте обработки нуклеоидного комплекса Е. coli ферментами, разрушающими белки и РНК, возникают открытые кольцевые формы хромосомной ДНК, которые изучают с помощью электронномикроскопических и радиобиологических методов (рис. 5.35). При этом можно определить длину молекул.

Рис. 5.35. Радиоавтограф кольцевой молекулы ДНК хромосомы Е. coli. Виден участок начавшейся репликации молекулы
Размеры хромосомного генома бактерий позволяют им иметь более значительное, чем у вирусов, количество структурных генов для синтеза белков, обеспечивающих все процессы жизнедеятельности этих организмов. Так, например, хромосомная ДНК клеток Е. coli содержит 4,2 х 10 б пар нуклеотидов, что было бы достаточным для формирования примерно 4 000 генов среднего размера. К настоящему времени в единственной кольцевой хромосоме (одной группе сцепления) этих бактерий идентифицировано более 650 разных генных локусов.
В хромосомах бактерий, вызывающих инфекционные заболевания животных и человека, обнаружены различные гены, детерминирующие патогенные свойства этих бактерий (формирование антигенных структур, синтез токсинов, способность разрушать эритроциты крови и др.). Значительный клинический интерес представляют также гены устойчивости бактерий к антибиотикам и другим антибактериальным препаратам.
Молекулярная биология, изучающая фундаментальные основы жизни, является в значительной степени детищем микробиологии. В качестве основных объектов изучения в ней используют вирусы и бактерии, а основное направление- молекулярная генетика основана на генетике бактерий и фагов.
Бактерии- удобный материал для генетики. Их отличает:
- относительная простота генома (сопокупности нуклеотидов хромосом);
- гаплоидность (один набор генов), исключающая доминантность признаков;
- различные интегрированные в хромосомы и обособленные фрагменты ДНК;
- половая дифференциация в виде донорских и реципиентных клеток;
- легкость культивирования, быстрота накопления биомасс.
Общие представления о генетике.
Ген- уникальная структурная единица наследственности, носитель и хранитель жизни. Он имеет три фундаментальные функции.
1.Непрерывность наследственности- обеспечивается механизмом репликации ДНК.
2.Управление структурами и функциями организма - обеспечивается с помощью единого генетического кода из четырех оснований (А- аденин, Т- тимин, Г- гуанин, Ц- цитозин). Код триплетный, поскольку кодон- функциональная единица, кодирующая аминокислоту, состоит из трех оснований (букв).
3.Эволюция организмов- благодаря мутациям и генетическим рекомбинациям.
В узкоспециальном плане ген чаще всего представляет структурную единицу ДНК, расположение кодонов в которой детерминирует первичную структуру соответствующей полипептидной цепи (белка). Хромосома состоит из особых функциональных единиц- оперонов.
Основные этапы развития (усложнения) генетической системы можно представить в виде следующей схемы:
кодон à ген à оперон à геном вирусов и плазмид à хромосома прокариот (нуклеоид) à хромосомы эукариот (ядро).
Генетический материал бактерий.
1.Ядерные структуры бактерий- хроматиновые тельца или нуклеоиды (хромосомная ДНК). У бактерий одна замкнутая кольцевидная хромосома (до 4 тысяч отдельных генов). Бактериальная клетка гаплоидна, а удвоение хромосомы (репликация ДНК) сопровождается делением клетки. Вегетативная репликация хромосомной (и плазмидной) ДНК обусловливает передачу генетической информации по вертикали- от родительской клетки- к дочерней. Передача генетической информации по горизонтали осуществляется различными механизмами- в результате конъюгации, трансдукции, трансформации, сексдукции.
2.Внехромосомные молекулы ДНК представлены плазмидами, мигрирующими генетическими элементами- транспозонами и инсервационными (вставочными) или IS- последовательностями.
Плазмиды- экстрахромосомный генетический материал (ДНК), более просто устроенные по сравнению с вирусами организмы, наделяющие бактерии дополнительными полезными свойствами. По молекулярной массе плазмиды значительно меньше хромосомной ДНК, содержат от 40 до 50 генов.
Их объединение в одно царство жизни с вирусами связано с наличием ряда общих свойств- отсутствием собственных систем мобилизации энергии и синтеза белка, саморепликацией генома, абсолютным внутриклеточным паразитизмом.
Их выделение в отдельный класс определяется существенными отличиями от вирусов.
1.Среда их обитания- только бактерии (среди вирусов , кроме вирусов бактерий- бактериофагов имеются вирусы растений и животных).
2.Плазмиды сосуществуют с бактериями, наделяя их дополнительными свойствами. У вирусов эти свойства могут быть только у умеренных фагов при лизогении бактерий, чаще же всего вирусы вызывают отрицательный последствия, лизис клеток.
3.Геном представлен двунитевой ДНК.
4.Плазмиды представляют собой “голые” геномы, не имеющие никакой оболочки, их репликация не требует синтеза структурных белков и процессов самосборки.
Плазмиды могут распространяться по вертикали (при клеточном делении) и по горизонтали, прежде всего путем конъюгационного переноса. В зависимости от наличия или отсутствия механизма самопереноса (его контролируют гены tra- оперона) выделяют конъюгативные и неконъюгативные плазмиды. Плазмиды могут встраиваться в хромосому бактерий- интегративные плазмиды или находиться в виде отдельной структуры- автономные плазмиды ( эписомы).
Классификация и биологическая роль плазмид.
Функциональная классификация плазмид основана на свойствах, которыми они наделяют бактерии. Среди них- способность продуцировать экзотоксины и ферменты, устойчивость к лекарственным препаратам, синтез бактериоцинов.
Основные категории плазмид.
1.F- плазмиды - донорские функции, индуцируют деление (от fertility - плодовитость). Интегрированные F - плазмиды- Hfr- плазмиды (высокой частоты рекомбинаций).
2.R- плазмиды (resistance) - устойчивость к лекарственным препаратам.
3.Col- плазмиды- синтез колицинов (бактериоцинов)- факторов конкуренции близкородственных бактерий (антогонизм). На этом свойстве основано колицинотипирование штаммов.
4.Hly- плазмиды- синтез гемолизинов.
5.Ent- плазмиды- синтез энтеротоксинов.
6.Tox- плазмиды- токсинообразование.
Близкородственные плазмиды не способны стабильно сосуществовать, что позволило объединить их по степени родства в Inc- группы (incompatibility- несовместимость).
Биологическая роль плазмид многообразна, в том числе:
- контроль генетического обмена бактерий;
- контроль синтеза факторов патогенности;
- совершенствование защиты бактерий.
Бактерии для плазмид- среда обитания, плазмиды для них- переносимые между ними дополнительные геномы с наборами генов, благоприятствующих сохранению бактерий в природе.
Мигрирующие генетические элементы - отдельные участки ДНК, способные определять свой перенос между хромосомами или хромосомой и плазмидой с помощью фермента рекомбинации транспозазы. Простейшим их типом являются инсерционные последовательности (IS- элементы) или вставочные элементы, несущие только один ген транспозазы, с помощью которой IS- элементы могут встраиваться в различные участки хромосомы. Их функции- координация взаимодействия плазмид, умеренных фагов, транспозонов и генофора для обеспечения репродукции, регуляция активности генов, индукция мутаций. Величина IS- элементов не превышает 1500 пар оснований.
Транспозоны (Tn- элементы) включают до 25 тысяч пар нуклеотидов, содержат фрагмент ДНК, несущий специфические гены, и два Is- элемента. Каждый транспозон содержит гены, привносящие важные для бактерии характеристики, как и плазмиды (множественная устойчивость к антибиотикам, токсинообразование и т.д.). Транспозоны- самоинтегрирующиеся фрагменты ДНК, могут встраиваться и перемещаться среди хромосом, плазмид, умеренных фагов, т.е. обладают потенциальной способностью распространяться среди различных видов бактерий.
Понятие о генотипе и фенотипе.
Генотип- вся совокупность имеющихся у организма генов.
Фенотип- совокупность реализованных (т.е. внешних) генетически детерминированных признаков, т.е. индивидуальное (в определенных условиях внешней среды) проявление генотипа. При изменении условий существования фенотип бактерий изменяется при сохранении генотипа.
Изменчивость у бактерий может быть ненаследуемой (модификационной) и генотипической (мутации, рекомбинации).
Временные, наследственно не закрепленные изменения, возникающие как адаптивные реакции бактерий на изменения окружающей среды, называются модификациями (чаще - морфологические и биохимические модификации). После устранения причины бактерии реверсируют к исходному фенотипу.
Стандартное проявление модификации- распределение однородной популяции на две или более двух типов- диссоциация. Пример- характер роста на питательных средах: S- (гладкие) колонии, R- (шероховатые) колонии, M- (мукоидные, слизистые) колонии, D- (карликовые) колонии. Диссоциация протекает обычно в направлении Sà R. Диссоциация сопровождается изменениями биохимических, морфологических, антигенных и вирулентных свойств возбудителей.
Мутации- скачкообразные изменения наследственного признака. Могут быть спонтанные и индуцированные, генные (изменения одного гена) и хромосомные (изменения двух или более двух участков хромосомы).
Одновременно у бактерий имеются различные механизмы репарации мутаций, в том числе с использованием ферментов- эндонуклеаз, лигаз, ДНК- полимеразы.
Генетические рекомбинации- изменчивость, связанная с обменом генетической информации. Генетические рекомбинации могут осуществляться путем трансформации, трансдукции, конъюгации, слияния протопластов.
1.Трансформация- захват и поглощение фрагментов чужой ДНК и образование на этой основе рекомбинанта.
2.Трансдукция- перенос генетического материала фагами (умеренными фагами- специфическая трансдукция).
3.Конъюгация- при непосредственном контакте клеток. Контролируется tra (transfer) опероном. Главную роль играют конъюгативные F- плазмиды.
Геном вирусов содержит или РНК, или ДНК (РНК- и ДНК- вирусы соответственно). Выделяют позитивную (+) РНК, обладающую матричной активностью и соответственно- инфекционными свойствами, и негативную ( - ) РНК, не проявляющую инфекционные свойства, которая для воспроизводства толжна транскрибироваться (превращаться) в +РНК. Механизмы репродукции различных вирусов очень сложные и существенно отличаются. Основные их схематические варианты представлены ниже.
1. вирионная (матричная) +РНК à комплементарная -РНК (в рибосомах) à вирионная +РНК.
2. - РНК à вирусная (информационная) +РНК à - РНК (формируется на геноме зараженной клетки).
3. однонитевая ДНК: +ДНК à +ДНК -ДНК à +ДНК -ДНК +ДНК à +ДНК.
4. ретровирусная однонитевая РНК: РНК à ДНК (провирус) à РНК.
5. двунитевая ДНК: разделение нитей ДНК и формирование на каждой комплементарной нити ДНК.
Генофонд вирусов создается и пополняется из четырех основных источников:
двух внутренних (мутации, рекомбинации) и двух внешних (включение в геном генетического материала клетки хозяина, поток генов из других вирусных популяций).
Комплементация- функциональное взаимодействие двух дефектных вирусов, способствующее их репликации и горизонтальной передаче.
Фенотипическое смешивание- при заражении клетки близкородственными вирусами с образованием вирионов с гибридными капсидами, кодируемыми геномами двух вирусов.
Популяционная изменчивость вирусов связана с двумя разнонаправленными процессами - мутациями и селекцией, связанными с внешней средой как индуктором мутаций и фактором стабилизирующего отбора. Гетерогенность вирусных популяций- адаптационный генетический механизм, способствующий пластичности (устойчивости, приспособляемости) популяций, фактор эволюции и сохранения видов во внешней среде.
Генофонд вирусных популяций сохраняется за счет нескольких механизмов:
- восстановления изменчивости за счет мутаций;
- резервирующих механизмов (возможность перехода любых, даже негативных мутаций в следующую генерацию)- комплементация, рекомбинация;
- буферных механизмов (образование дефектных вирусных частиц, иммунных комплексов и др.), способствующие сохранению вируса в изменяющихся внешних условиях.
Читайте также:
