
Дэвид балтимор классификация вирусов

Система классификации вирусов Балтимора была предложена в 1971 году Нобелевским лауреатом Дэвидом Балтимором.
Скачать:
| Вложение | Размер |
|---|---|
| klassifikatsiya_virusov_po_baltimoru.pptx | 1.98 МБ |
Подписи к слайдам:
Классификация вирусов по Балтимору .
Система классификации вирусов Балтимора была предложена в 1971 году Нобелевским лауреатом Дэвидом Балтимором. Классификация подразделяет вирусы на 7 групп в зависимости от механизма образования вирусной МРНК(молекулы матричной РНК ,по которой производится синтез белка) в клетке-хозяине.
Класс I: вирусы, содержащие двуцепочечную ДНК Для репликации попадают в ядро клетки, так как им требуется клеточная ДНК-полимераза. Не имеют РНК-стадии (к примеру, герпесвирусы , поксвирусы , паповавирусы , мимивирус ).
Класс II: вирусы, содержащие одноцепочечную молекулу ДНК. Вирусы реплицируют геномную ДНК в ядре и в ходе репликации образуют интермедиат — двуцепочечную ДНК . В этом случае ДНК всегда положительной полярности.(Вирусы семейств Circoviridae и Parvoviridae )
Класс III: вирусы, в которых РНК способна к репликации (редупликации) Вирусы реплицируют геномную РНК в цитоплазме и используют полимеразы хозяина в меньшей степени, чем ДНК-вирусы( Reoviridae и Birnaviridae )
Класс IV: Вирусы, содержащие одноцепочечную молекулу РНК положительной полярности. Непосредственно на (+) геномной РНК вирусов класса может идти синтез белка на рибосомах клетки хозяина. (представителей семейств Astroviridae , Caliciviridae , Coronaviridae , Flaviviridae , Picornaviridae , Arteriviridae и Togaviridae . )Классифицируются на 2 группы:вирусоы с полицистронной мРНК трансляция приводит к образованию полипротеина , вирусы со сложной трансляцией — синтез белка идет со сдвигом рамки считывания.
Класс V: Вирусы, содержащие одноцепочечную молекулу РНК негативной или двойной полярности Геномные РНК вирусов класса V не могут быть транслированы на рибосомах клетки хозяина, предварительно требуется транскрипция вирусными РНК-полимеразами в (+)РНК.(семейства Arenaviridae , Orthomyxoviridae , Paramyxoviridae , Bunyaviridae , Filoviridae, Rhabdoviridae и Deltavirus . ) классифицируют на две группы : вирусы, содержащие несегментированный геном, вирусы с сегментированными геномами.
Н а первом этапе репликации происходит транскрипция (−)РНК вирусной РНК-зависимой РНК-полимеразой в моноцистронную мРНК , и далее синтезируются дополнительные копии (+)РНК, служащие матрицами для синтеза геномных (−)РНК. Репликация геномных РНК таких вирусов осуществляется в цитоплазме. Вирусы, содержащие несегментированный геном.
Вирусы с сегментированными геномами. Репликация геномных РНК которых происходит в клеточном ядре, вирусная РНК-зависимая РНК-полимераза синтезирует моноцистронные мРНК с каждого сегмента генома. Наибольшим отличием данной группы вирусов от другой группы пятого класса состоит в том, что репликация осуществляется в двух местах.
Класс VI: Вирусы, содержащие одноцепочечную (+)РНК, реплицирующиеся через стадию ДНК. Вирусы класса используют фермент обратную транскриптазу для превращения (+)РНК в ДНК. Вместо использования РНК в качестве матрицы для синтеза белков, вирусы этого класса используют матрицу ДНК, которая встраивается в геном хозяина ферментом интегразой . Дальнейшая репликация происходит при помощи полимераз клетки хозяина.( Ретровирусы (например, ВИЧ))
Класс VII: вирусы, содержащие двуцепочечную ДНК, реплицирующиеся через стадию одноцепочечной РНК Вирусы имеют двуцепочечную геномную ДНК, которая ковалентно замкнута в форме кольца и является матрицей для синтеза мРНК вируса, а также субгеномных РНК. Субгеномная РНК служит матрицей для синтеза ДНК-генома ферментом обратной транскриптазой вируса(семейство Hepadnaviridae (например ,гепатит В)
Вирусы, содержащие двуцепочечную ДНК для репликации попадают в ядро клетки, так как им требуется клеточная ДНК-полимераза. Также репликация ДНК этих вирусов сильно зависит от стадии клеточного цикла. В некоторых случаях вирус может вызывать деления клетки, что может приводить к раковому перерождению. Примерами таких вирусов являются Herpesviridae, Adenoviridae и Papovaviridae.
У представителей семейства Poxvirus геномная ДНК реплицируется не в ядре.
Вирусы семейств Circoviridae и Parvoviridae реплицируют геномную ДНК в ядре и в ходе репликации образуют интермедиат — двуцепочечную ДНК.
Как и большинство РНК-вирусов, представители класса III реплицируют геномную РНК в цитоплазме и используют полимеразы хозяина в меньшей степени, чем ДНК-вирусы. Класс III включает в себя два крупных семейства Reoviridae и Birnaviridae. Репликация моноцистронная, геном сегментирован, каждый ген кодирует один белок.
Классы IV и V включают вирусы двух типов, репликация которых не зависит от стадии клеточного цикла. Наряду с вирусами, содержащими двуцепочечную ДНК, эти вирусы наиболее изучены.
Непосредственно на (+) геномной РНК вирусов IV класса может идти синтез белка на рибосомах клетки хозяина. Вирусы классифицируют на две группы, в зависимости от особенностей РНК:
- у вирусов с полицистронной мРНК трансляция приводит к образованию полипротеина, который затем разрезается на зрелые белки. С одной цепи РНК может синтезироваться несколько разных белков, что снижает длину генов.
- вирусы со сложной транскрипцией содержат субгеномные мРНК, синтез белка идет со сдвигом рамки считывания, также используется протеолитический процессинг полипротеинов. Эти механизмы обеспечивают синтез разных белков с одной цепи РНК.
Примеры вирусов данного класса включают представителей семейств Astroviridae, Caliciviridae, Coronaviridae, Flaviviridae, Picornaviridae, Arteriviridae и Togaviridae.
Геномные РНК вирусов класса V не могут быть транслированы на рибосомах клетки хозяина, предварительно требуется транскрипция вирусными РНК-полимеразами в (+)РНК. Вирусы пятого класса классификации по Балтимору классифицируют на две группы:
- вирусы, содержащие несегментированный геном, на первом этапе репликации происходит транскрипция (−)РНК вирусной РНК-зависимой РНК-полимеразной в моноцистронную мРНК, и далее синтезируются дополнительные копии (+)РНК, служащие матрицами для синтеза геномных (−)РНК. Репликация геномных РНК таких вирусов осуществляется в цитоплазме.
- вирусы с сегментированными геномами, репликация геномных РНК которых происходит в клеточном ядре, вирусная РНК-зависимая РНК-полимераза синтезирует моноцистронные мРНК с каждого сегмента генома. Наибольшим отличием данной группы вирусов от другой группы пятого класса состоит в том, что репликация осуществляется в двух местах.
Представители данного класса входят в состав семейств: Arenaviridae, Orthomyxoviridae, Paramyxoviridae, Bunyaviridae, Filoviridae и Rhabdoviridae.
Наиболее хорошо изученным семейством данного класса вирусов, являются ретровирусы. Вирусы класса VI используют фермент обратную транскриптазу для превращения (+)РНК в ДНК. Вместо использования РНК в качестве матрицы для синтеза белков, вирусы этого класса используют матрицу ДНК, которая встраивается в геном хозяина ферментом интегразой. Дальнейшая репликация происходит при помощи полимераз клетки хозяина. Наиболее хорошо изученным представителем данной группы вирусов является ВИЧ.
Небольшая группа вирусов, в состав которой входит вирус гепатита В, представитель семейства Hepadnaviridae, имеют двуцепочечную геномную ДНК, которая ковалентно замкнута в форме кольца и является матрицей для синтеза мРНК вируса, а также субгеномных РНК. Субгеномная РНК служит матрицей для синтеза ДНК-генома ферментом обратной транскриптазой вируса.
Классификации Балтимор , разработанные Дэвид Балтимор , является классификация вирусов системы , что группы вирусов в семьи, в зависимости от их типа генома ( ДНК , РНК , одноцепочечные (сс), двухцепочечный (DS), и т.д ..) и их метод репликации .
содержание
классификации
Классифицируя вирусы в соответствии с их геномом означает, что те, в данной категории будет все ведут себя почти так же путь, который предлагает некоторое представление о том, как приступить к дальнейшим исследованиям. Короче:
Эти типы вирусов необходимо ввести хост - ядро , прежде чем они способны к репликации. Кроме того, эти вирусы требуют клеток - хозяин , полимераз для репликации вируса генома , и, следовательно, в значительной степени зависит от клеточного цикла . Правильная инфекция и производство потомства требует, чтобы электролизер в репликации, так как она во время репликации , что полимераза клетки активна. Вирус может вызвать клетку принудительно проходить деление клеток , что может привести к трансформации клетки и, в конечном счете, рак . Примеры включают герпесвирусов , Adenoviridae и Papovaviridae .
Существует только один хорошо изученный пример , в котором вирус класс-не реплицируется в ядре: поксвирусных семья, высоко патогенный вирус , который заражает позвоночные и включает оспу вирус.
МРНК регулярно транскрибируется с вирусной ДНК с помощью хозяина РНК-полимеразы II. Это производит два вида мРНК: 1) в начале мРНК, переписана до синтеза вирусной ДНК, и 2) в конце мРНК, переписана из потомства ДНК.
Вирусы в этой категории включают Anelloviridae , Circoviridae и Parvoviridae (которые инфицируют позвоночных животных), в Geminiviridae и Nanoviridae (которые заражают растения), и Microviridae (которые заражают прокариоты ). Большинство из них имеет круговые геном (парвовирусы являются единственным известным исключением). Эукариоты-заражать репликации вирусов в основном в пределах ядра - обычно через прокатку механизма окружности , образуя двухцепочечные ДНК промежуточных в процессе. Превалирующий но бессимптомно человек Anellovirus, называемое Переливание передающийся Вирус (TTV), входят в этой классификации.
Вирусы оцРНК принадлежат к классу IV или V классификации Baltimore. Они могут быть сгруппированы в положительном смысле или отрицательном смысле по смыслу полярности РНК. Одноцепочечной РНК является общим признаком этих вирусов. Репликация вирусов происходит в цитоплазме или ядре (для сегментированных вирусов класса V). Класс IV и V оцРНК вирусы не зависит так сильно, как ДНК-вирусов на клеточном цикле.
РНК - вирусы с положительным смыслом и вообще все РНК , определенные как положительный смысл могут быть доступны непосредственно принимающими рибосомами немедленно образуют белки. Их можно разделить на две группы, каждая из которых размножаются в цитоплазме:
- Вирусы с полицистронныммРНКом , где РНК - геном образует мРНК и транслируется в полипротеиновом продукт , который затем расщепляется с образованием зрелых белков. Это означает , что ген может использовать несколько методов , в которых для производства белков из одной и той же цепи РНК, все в целях уменьшения размера его гена.
- Вирусы со сложной транскрипцией, для которых субгеномного мРНК, рибосомная frameshifting и протеолитическая могут быть использована обработка полипротеинов. Все из которых являются различные механизмы , с помощью которых для получения белков из одной и той же цепи РНК.
- Вирусы , содержащие нерасчлененными геномы , для которых первым шагом в репликации транскрипции от (-) - многожильного генома вирусной РНК-зависимой РНК - полимеразы с получением monocistronic мРНК , которые кодируют различные вирусные белки. Геном копия положительного смысла затем производится , который служит в качестве матрицы для производства (-) - нити генома. Репликация в цитоплазме.
- Вирусы с сегментированных геномов , для которых репликация происходит в ядре и для которых вирусной РНК-зависимой РНК - полимеразы производит monocistronic мРНК из каждого сегмента генома. Самая большая разница между этими двумя местом репликации.
Примеры этого класса включают в семьях Arenaviridae , Orthomyxoviridae , Paramyxoviridae , буньявирусы , Filoviridae и Rhabdoviridae (последний из которых включает в себя вирус бешенства ).
Хорошо изученное семейство этого класса вирусов включают ретровирусы . Характерный признак является использование обратной транскриптазы для преобразования РНКА положительных смыслового в ДНК. Вместо того , чтобы использовать РНК для шаблонов белков, они используют ДНК для создания шаблонов, который сращивание в геном хозяина с помощью интегразы . Репликация может затем начать с помощью полимераз принимающей клетки.
Эта небольшая группа вирусов, примером которых может служить гепатита вируса (который находится в Hepadnaviridae семьи), имеют двухцепочечной, гэп геном , который затем заполняют с образованием ковалентно замкнутый круг ( ковалентно замкнутую кольцевую ДНК ) , который служит в качестве шаблона для производства вирусных мРНК и субгеномного РНК. РНК pregenome служит шаблоном для обратной транскриптазы вируса для производства генома ДНК.
вирусолог, биохимик и молекулярный биолог, член Национальной АН США
Настоящее имя: Baltimore David Год рождения: 1938 Годы жизни: 1938 — н/а Место рождения: г. Нью-Йорк, США Место смерти: Разделы каталога: Биологические науки Категория: Биохимия · Вирусология · Молекулярная биология -->
Балтимор Дейвид (Baltimore David) - американский вирусолог, биохимик и молекулярный биолог, член Национальной АН США. В 1970 выделил (одновременно с Х. Темином и независимо от него) из опухолеродного РНК-содержащего вируса фермент РНК-зависимую ДНК-полимеразу (ревертазу). Показал, что генетическая информация опухолеродного РНК-содержащего вируса с помощью этого фермента путём обратной транскрипции переписывается в последовательность ДНК-продукта, который интегрируется в геном клетки, в результате чего нормальная клетка превращается в раковую. Нобелевская премия (1975), совместно с Р. Дульбекко и Х. Темином.
Дейвид Балтимор родился в Нью-Йорке. Начальное и среднее образование Дейвид получил в государственной школе, где проявилась его склонность к биологии и математике. По программе для одаренных учеников средней школы он провел лето в Джексоновской лаборатории исследования генетики млекопитающих в Бар-Харборе (штат Мэн). Здесь он познакомился со студентом-биологом Ховардом Темином.
В 1956 г. Балтимор закончил среднюю школу и поступил в колледж в Суортморе, штат Пенсильвания, где специализировался по химии. В 1960 году получил диплом бакалавра с отличием и поступил в аспирантуру Массачусетского технологического института (МТИ) в Кембридже. Спустя год перешел в Рокфеллеровский университет в Нью-Йорке, где работал под руководством Ричарда Фрэнклина, признанного авторитета в вирусологии. В 1963/64 учебном году Балтимор продолжал исследования в постдокторантуре факультета биологии МТИ, потом работал в Нью-йоркском медицинском колледже Альберта Эйнштейна.
С 1965 по 1968 г. Д.Балтимор работал научным сотрудником в Институте биологических исследований им. Дж. Солка в Ла-Джолле (штат Калифорния). В институте он встретился с Ренато Дульбекко, разработавшим количественные методы экспериментального изучения генетики вирусов и классификацию различий между нормальными клетками и клетками, ставшими опухолевыми в результате вирусного заражения.
Затем Балтимор обратился к опухолеродным вирусам, многие из которых также содержали РНК, но не содержали ДНК. В серии экспериментов конца 1960-х – начала 1970-х годов Балтимор установил, что способность РНК-содержащих вирусов поражать клетки тканей человека и животных — это следствие так называемой обратной транскрипции, то есть возможности передачи генетической информации не от ДНК к РНК и затем к белку (транскрипция), а, наоборот, от молекулы РНК к ДНК. В 1970 г. Д.Балтимор и Х.Темин независимо друг от друга выделили РНК-зависимую ДНК-полимеразу. В мае того же года Х.Темин сообщил о своем открытии на Х Международном конгрессе Международного союза борьбы с раком, а затем Балтимор доложил о своих результатах на симпозиуме в Колд Спринг Харборе. Оба исследователя опубликовали свои данные в июне 1970 г. Вскоре они были подтверждены Соломоном Шпигельманом, директором Научно-исследовательского института рака Колумбийского университета. В течение последующих 10 лет процесс обратной транскрипции стал одной из центральных тем исследований во всем мире.
В 1973 г. Балтимор был назначен на должность профессора микробиологии, учрежденную Американским онкологическим обществом, в МТИ. Продолжая исследования по изучению фермента обратной транскриптазы у других онкогенных вирусов, Балтимор обнаружил восемь вирусов, обладающих данным ферментом.
Работая в МТИ, в настоящее время Балтимор также является консультантом по медицине и педиатрической онкологии в Детском госпитальном медицинском центре и Институте рака Сиднея Фарбера в Бостоне (штат Массачусетс). В 1986 г. он был председателем комитета Национальной академии наук США по изучению СПИДа.
Балтимор является членом многих профессиональных обществ, включая Американскую ассоциацию содействия развитию науки, Американскую академию наук и искусств и Национальную академию наук США. Кроме Нобелевской премии, он удостоен премии Эли Лилли за исследования в области микробиологии и иммунологии Американского микробиологического общества (1971), международной награды Гарднеровского фонда (1974) и премии Национальной академии наук американского Фонда Стила за исследования в области молекулярной биологии (1974).
Балтимор – сторонник осторожного применения методик генной инженерии, поэтому он присоединился к группе специалистов по молекулярной биологии, предложивших объявить мораторий на некоторые виды экспериментов с ДНК.
Эта статья еще не написана, но вы можете сделать это.
Часть I. Общая молекулярная биология вирусов.
1. Классификация вирусов по Балтимору. Классификация вирусов МКТВ (ICTV). Критерии выделения таксонов. 2
2. Стратегии репликации и экспрессии вирусных геномов. Варианты репликации ДНК-вирусов. Жизненные циклы (+) и (-) РНК вирусов. Ретро- и параретровирусы. 5
3. Тактические приёмы, используемые вирусами при экспрессии геномов. Сдвиг рамки считывания, протечка при сканировании, внутренняя инициация трансляции, рибосомальный пропуск, рибосомальное шунтирование. 11
4. Общие принципы структурной организации вирусных капсидов. Виды симметрии капсидов. Функции капсидной оболочки. Примеры вирусов с различными капсидами. 12
5. Принцип строения ИК. Ассиметричная ячейка. Капсомеры. Число Т. Примеры вирусов с ИК (смотри предыдущий вопрос!). 16
6. ИК с Т=7, состоящий из пентонов. Капсиды с различными белками в ассиметрической ячейке (Т=pseudo3). Двойные капсиды. 17
7. Мимивирус. Особенности строения. Вирус Sputnik. 18
8. Капсиды со спиральной симметрией, общие принципы. Вирус табачной мозаики. Способы укладки (фолды) вирусных структурных белков. 19
9. Капсиды вирусов с (-)РНК-геномом. Организация капсидов Филовирусов, Рабдовирусов, Орто- и Парамиксовирусов. 22
10. Капсиды бактериофагов. Семейства Siphoviridae, Podoviridae и Myoviridae. Семейство Inoviridae (нитчатые фаги). 25
11. Сборка фага Т4. Особенности, основные этапы сборки. 30
12. Механизмы проникновения вирусов в клетку. Фузия, эндоцитоз (клатрин-зависимый/кавеолин-зависимый), проникновение с образованием поры. 32
13. Макропиноцитоз. Варианты проникновения вируса из эндосомы в цитоплазму. Транспорт вирусных частиц по клетке. 39
14. Вход и выход вирусов из ядра. Варианты выхода вирусов из клетки. Вирусные фабрики. 43
15. Особенности врождённого и адаптивного иммунитетов. Toll-подобные рецепторы. Дефензины. Интерфероны. NK-клетки. 46
16. Молекулы главного комплекса гистосовместимости (MHC). Т-клеточные рецепторы (TCR), двойное распознавание антигена. В-клеточные рецепторы (BCR). 51
17. Вакцинация. Живые и инактивированные вирусные вакцины. Субъединичные вакцины. Примеры вакцин. 55
Классификация вирусов по Балтимору. Классификация вирусов МКТВ (ICTV). Критерии выделения таксонов.
1. Классификация вирусов по Балтимору — классификация вирусов в группы в зависимости от типа геномной нуклеиновой кислоты (ДНК, РНК, одноцепочечная, двуцепочечная) и способа репликации. Предложена американским учёным Дэвидом Балтимором в 1971 году.
Класс I: вирусы, содержащие дц ДНК
Вирусы, содержащие двуцепочечную ДНК для репликации попадают в ядро клетки, так как им требуется клеточная ДНК-полимераза. Также репликация ДНК этих вирусов сильно зависит от стадии клеточного цикла. В некоторых случаях вирус может вызывать деления клетки, что может приводить к раковому перерождению. Примерами таких вирусов являются Herpesviridae, Adenoviridae и Papovaviridae.
У представителей семейства Poxvirus геномная ДНК реплицируется не в ядре.
Класс II: оц ДНК
Вирусы семейств Circoviridae и Parvoviridae реплицируют геномную ДНК в ядре и в ходе репликации образуют интермедиат — двуцепочечную ДНК.
Класс III: дц РНК
Как и большинство РНК-вирусов, представители класса III реплицируют геномную РНК в цитоплазме и используют полимеразы хозяина в меньшей степени, чем ДНК-вирусы. Класс III включает в себя два крупных семейства — Reoviridae и Birnaviridae. Репликация моноцистронная, геном сегментирован, каждый ген кодирует один белок.
Классы IV и V: оц РНК
Классы IV и V включают вирусы двух типов, репликация которых не зависит от стадии клеточного цикла. Наряду с вирусами, содержащими двуцепочечную ДНК, эти вирусы наиболее изучены.
Класс IV: оц (+)РНК
Непосредственно на (+) геномной РНК вирусов IV класса может идти синтез белка на рибосомах клетки хозяина. Вирусы классифицируют на две группы, в зависимости от особенностей РНК:
· у вирусов с полицистронной мРНК трансляция приводит к образованию полипротеина, который затем разрезается на зрелые белки. С одной цепи РНК может синтезироваться несколько разных белков, что снижает длину генов.
· вирусы со сложной трансляцией — синтез белка идет со сдвигом рамки считывания, также используется протеолитический процессинг полипротеинов. Эти механизмы обеспечивают синтез разных белков с одной цепи РНК.
Примеры вирусов данного класса включают представителей семейств Astroviridae, Caliciviridae, Coronaviridae, Flaviviridae, Picornaviridae, Arteriviridae иTogaviridae.
Класс V: оц (−)РНК
Геномные РНК вирусов класса V не могут быть транслированы на рибосомах клетки хозяина, предварительно требуется транскрипция вирусными РНК-полимеразами в (+)РНК. Вирусы пятого класса классификации по Балтимору классифицируют на две группы:
· вирусы, содержащие несегментированный геном, на первом этапе репликации происходит транскрипция (−)РНК вирусной РНК-зависимой РНК-полимеразой в моноцистронную мРНК, и далее синтезируются дополнительные копии (+)РНК, служащие матрицами для синтеза геномных (−)РНК. Репликация геномных РНК таких вирусов осуществляется в цитоплазме.
· вирусы с сегментированными геномами, репликация геномных РНК которых происходит в клеточном ядре, вирусная РНК-зависимая РНК-полимераза синтезирует моноцистронные мРНК с каждого сегмента генома. Наибольшим отличием данной группы вирусов от другой группы пятого класса состоит в том, что репликация осуществляется в двух местах.
Представители данного класса входят в состав семейств: Arenaviridae, Orthomyxoviridae, Paramyxoviridae, Bunyaviridae, Filoviridae, Rhabdoviridae и Deltavirus.
Класс VI: оц (+)РНК, реплицирующиеся через стадию ДНК
Наиболее хорошо изученным семейством данного класса вирусов, являются ретровирусы. Вирусы класса VI используют фермент обратную транскриптазудля превращения (+)РНК в ДНК. Вместо использования РНК в качестве матрицы для синтеза белков, вирусы этого класса используют матрицу ДНК, которая встраивается в геном хозяина ферментом интегразой. Дальнейшая репликация происходит при помощи полимераз клетки хозяина. Наиболее хорошо изученным представителем данной группы вирусов является ВИЧ.
Класс VII: дц ДНК, реплицирующиеся через стадию одноцепочечной РНК
Небольшая группа вирусов, в состав которой входит вирус гепатита B, представитель семейства Hepadnaviridae, имеют двуцепочечную геномную ДНК, которая ковалентно замкнута в форме кольца и является матрицей для синтеза мРНК вируса, а также субгеномных РНК. Субгеномная РНК служит матрицей для синтеза ДНК-генома ферментом обратной транскриптазой вируса.
МКТВ (ICTV) –Международным Комитетом по Таксономии Вирусов в 1966 году была принята система классификации вирусов основанная на различии типа (РНК и ДНК), количества молекул нуклеотических кислот (одно- и двух-цепочечные) и на наличии или отсутствии оболочки ядра. Система классификации представляет собой серию иерархичных таксонов.
Принципы: выделяют Семейство (viridae), Род (virus), Вид (вир. Чумы плотоядных). Есть попытки объединить вирусы в большие таксоны Порядки (virales). Доп. Таксоны: подсемейство, штамм
Критерии: Семейство ( репродуктивная стратегия, структура вириона, структура генома), Род ( размер и организация генома, путь инфекции, гомологичные последовательности в геном), Вид ( серологическая кроссреактивность, круг хозяев, тканевой тропизм, патогенез, географич. Распр.
Международный комитет по таксономии вирусов – МКТВ (International committee on taxonomy of viruses - ICTV). (Доклад опубликован в 2011 году. Определено более 2200 видов, 349 родов, 19 подсемейств, 87 семейств и 6 порядков.)
Вирусный вид – политипическая категория (класс) вирусов, которая составляет реплицирующуюся линию и занимает особую экологическую нишу.
Штамм- отдельный изолят, происходящий из конкретного источника, как правило, характеризующегося по месту и времени, и имеющий собственное обозначение. Штаммы вируса могут пассироваться на восприимчивых животных или культурах ткани, сохраняя при этом свое обозначение. Если в процессе пассирования происходит изменение исходных свойств изолята, то измененный штамм получает название варианта, или деривата, и дополнительное обозначение. Часто штаммы имеют различия по биологическим свойствам, но не настолько выраженные, чтобы изменить их принадлежность к определенному типу вируса. Наиболее точно штамм вируса характеризует нуклеотидная последовательность (sequence) его генома, возможность определять которую появилась в последнее время.
Классификация вирусов
1. Нуклеиновая кислота
· одно или двуцепочечная
· сегментированны геном или нет
· линейный или кольцевой
· если РНК – какой полярности
2. Структура вириона
- икосаэндрическая, спиральная или комплексная симметрия
-наличие липопротеидной оболочки
Вирусы классифицируют по порядкам, семействам, родам и видам.
Порядки имеют латинское название с суффиксом -virales, семейства и подсемейства -viridae и -virinae, название родов заканчивается на -virus. В 6 порядков сгруппированы далеко не все вирусы.
3. Классификация вирусов в зависимости от генетического материала:
· Вирусы, содержащие двунитевую ДНК
· Вирусы, содержащие однонитевую ДНК
· Вирусы, содержащие двунитевую РНК
· Вирусы,содержащие однунитевую (+) РНК
· Вирусы, содержащие однунитевую (-) РНК
· РНК-содержащие вирусы, жизненный цикл которых включает стадию обратной транскрипции
· ДНК-содержащие вирусы, жизненный цикл которых включает стадию обратной транскрипции
· Субвирусные агенты (вироиды, вирусы-сателлиты, сателлитные нуклеиновые кислоты и прионы).
Критерии токсонов:
1) Тип нуклеиновой кислоты (ДНК или РНК) и ее структура (количество и форма цепей);
2) Стратегия вирусного генома;
3) Размер и морфология вириона, тип симметрии, число капсомеров;
4) Физико-химические свойства (устойчивость к температурам, рН, жирорастворителям и т.д.)
5) Наличие липопротеидной оболочки;
6) Антигенные свойства;
7) Патогенность, в т.ч. цитопатические изменения в клетках и внутриклеточные включения;
8) Круг восприимчивых хозяев;
9) Способ передачи;
10) Географическое распространение.
Строение вирусной частицы. Нуклеиновая кислота:
v дцДНК – вирусы
v оцДНК – вирусы
Ø РНК – содержащие вирусы
v дцРНК – вирусы
v (+)оцРНК – вирусы
v (-)оцРНК – вирусы
Ø Вирусы с обратной транскрипцией
МКТВ – Международный комитет по таксономии вирусов (1966)
Таксоны вирусов:
Внутри вида могут быть уровни:
Критерии семейства:
Семейство – группа родов вирусов, имеющих определенные общие признаки. Семейство обозначается одним словом на английском языке с окончанием viridae.
v Тип нуклеиновой кислоты (ДНК или РНК) и ее структура – количество цепей: односпиральная или двуспиральная; – форма цепей: линейная или кольцевая; непрерывная или сегментированная.
v Стратегия вирусного генома (способ репродукции по классификации Балтимора).
Критерии рода:
Род – это группа видов вирусов, имеющих определенные общие признаки, обозначается одним словом на английском языке с окончанием virus.
v Размер и морфология вириона, тип симметрии, число капсомеров.
v Наличие липопротеидной оболочки.
v Физико-химические свойства (устойчивость к температурам, рН среды, жирорастворителям и т.д.).
Критерии вида:
Ø Вид– это политетический класс вирусов (1), составляющих реплицирующуюся линию (2) и занимающих определенную экологическую нишу (3). Название должно состоять из min слов, сост. не только из слов хозяина и слова вирус, должно обозначаться на русском или англ. языке.
1) этот класс создают вирусы, имеющие ряд одинаковых признаков, среди которых нет единого общего признака.
2) вид образуют вирусы, имеющие общего предшественника
3) означает совокупность, т.н. биотических характеристик вирусов, к которым относится спектр восприимчивых хозяев, тканевой тропизм, вирулентность, патогенез, ответ со стороны хозяина, переносчики, среда обитания и т.д.
Критерии:
v Антигенные свойства;
v Патогенность, в т.ч. ЦПД в клетках и внутриклеточные включения;
v Круг восприимчивых хозяев;
v Способ передачи;
v Географическое распространение
. Подсемейство – обозначается одним словом на английском языке с окончанием virinae.
. Порядок – virales→
Вироиды классифицируются как вирусы. Рекомбинантные вирусы как виды или подвиды. Вирусы - сателлиты не классифицируются
2. Стратегии репликации и экспрессии вирусных геномов. Варианты репликации ДНК-вирусов. Жизненные циклы (+) и (-) РНК вирусов. Ретро- и параретровирусы.
+РНК вирусы.
Ø Голая (неочищенная) (+)РНК – инфекционная, для экспрессии генома не нужны вирусные белки.
Ø Жизненный цикл протекает в цитоплазме.
Ø В большинстве случаев геномная мРНК транслируется в полипротеин, который нарезается вирусными или клеточными протеазами на отдельные белки.
Ø Некоторые вирусы используют стратегию субгеномных РНК.

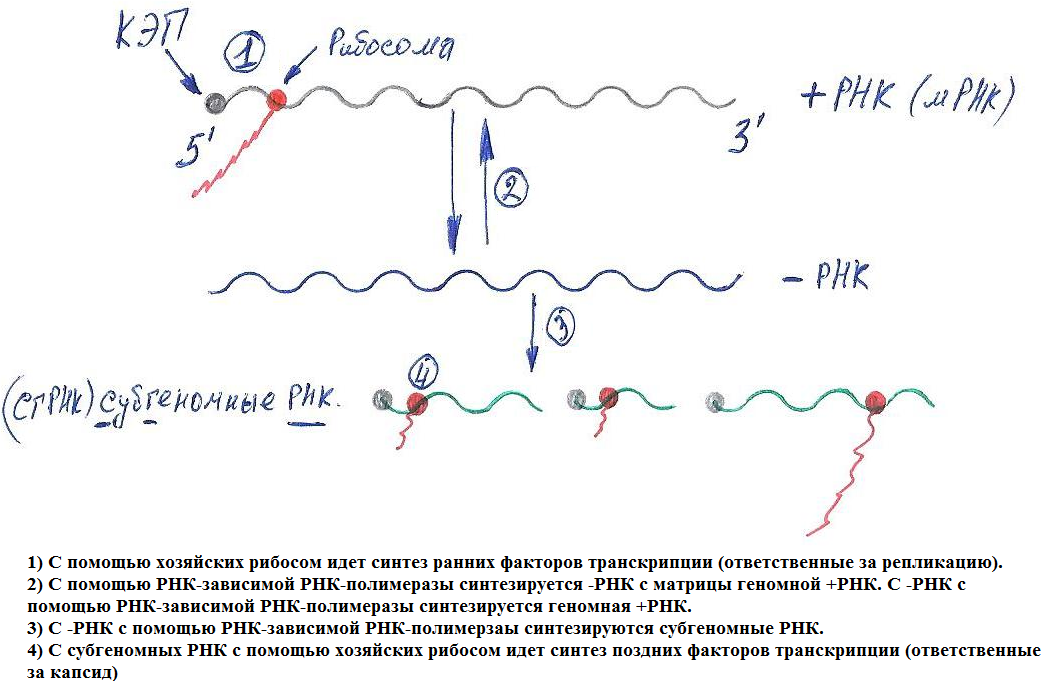
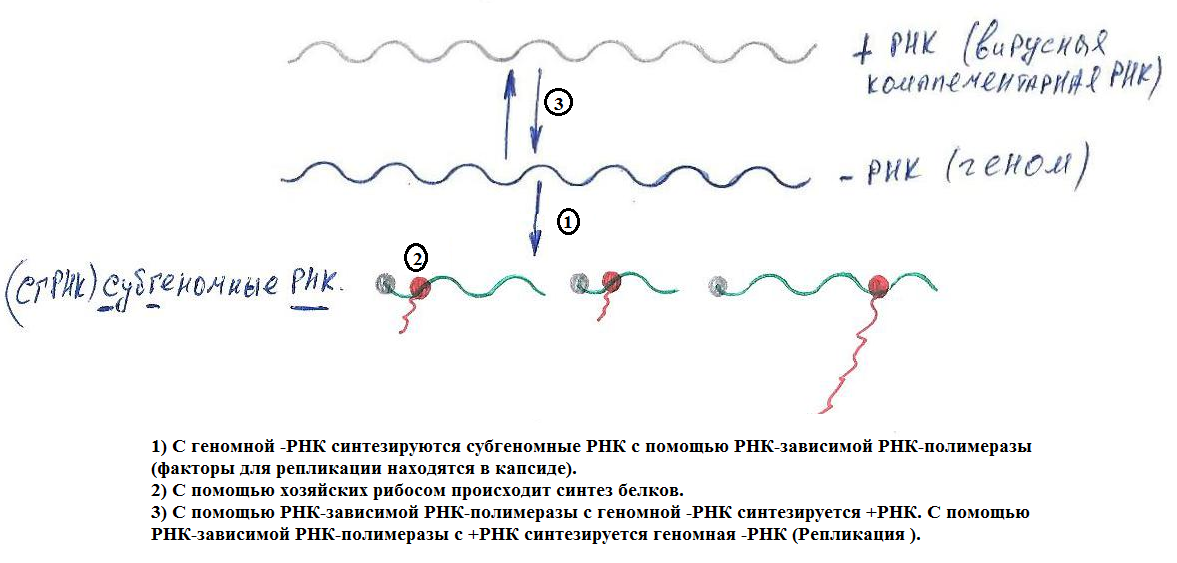
(-)РНК и (±)РНК – вирусы
Ø Для экспрессии генома необходима вирусная РНК-полимераза (присутствует в капсиде).
Ø Жизненный цикл протекает в цитоплазме (за искл. ортомиксовирусов и борнавирусов).
Ø Для всех капсидов (-)РНК – вирусов характерен спиральный тип симметрии и практически все имеют внешнюю липопротеидную мембрану.
Ø мРНК обычно моноцистроны.
Ø Антигеномные (комплементарные геному) РНК (вкРНК), несмотря на (+) – полярность не могут транслироваться.
Ø У амбиполярных (двуспиральные) – РНК – вирусов с антигенной РНК могут транскрибироваться мРНК (способность временного регулирования экспрессии генов).
На дцРНК запускается иммунный ответ, дцРНК – характерная стадия для всех РНК – вирусов.
Многие вирусы кодируют белки, блокирующие запуск иммунного ответа.

Дата добавления: 2018-06-01 ; просмотров: 538 ;
Читайте также:
