
Антигенная структура изменение антигенных свойств и плюрализм вирусов
В структуре вирусной частицы различают несколько групп антигенов: ядерные (или коровые), капсидные (или оболочечные) и суперкапсидные. На поверхности некоторых вирусных частиц встречаются особые V-антигены — гемагглютинин и фермент нейраминидаза. Антигены вирусов различаются по происхождению. Часть из них — вирусоспецифические. Информация об их строении картирована в нуклеиновой кислоте вируса. Другие антигены вирусов являются компонентами клетки хозяина (углеводы, липиды),] они захватываются во внешнюю оболочку вируса при его рождении путем почкования.
Антигенный состав вириона зависит от строения самой вирусной частицы. Антигенная специфичность простоорганизованных вирусов связана с рибо- и дезоксирибонуклеопротеинами. Эти вещества хорошо растворяются I в воде и поэтому обозначаются как S-антигены (от лат. solutio — раствор). У сложноорга- низованных вирусов часть антигена связанаснуклеокапсидом, а другая — локализуется во внешней оболочке — суперкапсиде.
Антигены многих вирусов отличаются высокой степенью изменчивости. Это связано с постоянным мутационным процессом, который претерпевает генетический аппарат вирусной частицы. Примером могут служить вирус гриппа, вирусы иммунодефицитов человека.
60. Учение об иммунитете. Определение и сущность понятия "иммунитет". Основные формы иммунного ответа.
Иммунитет – это способ защиты организма от генетически чужеродных веществ – антигенов экзогенного и эндогенного происхождения, направленный на поддержание и сохранение гомеостаза, структурной и функциональной целостности организма, биологической (антигенной) индивидуальности каждого организма и вида в целом.
Виды иммунного ответа
Иммунный ответ представляет собой реакцию организма на внедрение в него микробов или различных ядов. В целом, любое вещество, чья структура отличается от структуры тканей человека способно вызвать иммунный ответ. Исходя из механизмов, задействованных в его реализации, иммунный ответ может быть различным.
Во-первых, различаем специфический и неспецифический иммунный ответ.
Неспецифический иммунный ответ - это первый этап борьбы с инфекцией он запускается сразу же после попадания микроба в наш организм. В его реализации задействованы система комплимента, лизоцим, тканевые макрофаги. Неспецифический иммунный ответ практически одинаков для всех типов микробов и подразумевает первичное разрушение микроба и формирование очага воспаления. Воспалительная реакция это универсальный защитный процесс, который направлен на предотвращение распространения микроба. Неспецифический иммунитет определяет общую сопротивляемость организма. Люди с ослабленным иммунитетом чаще болеют различными заболеваниями.
Специфический иммунитет это вторая фаза защитной реакции организма. Основной характеристикой специфического иммунного ответа является распознавание микроба и выработка факторов защиты направленных специально против него. Процессы неспецифического и специфического иммунного ответа пересекаются и во многом дополняют друг друга. Во время неспецифического иммунного ответа часть микробов разрушается, а их части выставляются на поверхности клеток (например, макрофагов). Во второй фазе иммунного ответа клетки иммунной системы (лимфоциты) распознают части микробов, выставленные на мембране других клеток, и запускают специфический иммунный ответ как таковой.
Специфический иммунный ответ может быть двух типов: клеточный и гуморальный.
Клеточный иммунный ответ подразумевает формирование клона лимфоцитов (К-лимфоциты, цитотоксические лимфоциты), способных разрушать клетки мишени, мембраны которых содержат чужеродные материалы (например, вирусные белки).
Клеточный иммунитет задействован в ликвидации вирусной инфекции, а также таких типов бактериальных инфекций как туберкулез, проказа, риносклерома. Раковые клетки тоже разрушаются активированными лимфоцитами.
Гуморальный иммунный ответ опосредован В-лимфоцитами, которые после распознания микроба начинают активно синтезировать антитела по принципу один тип антигена – один тип антитела. На поверхности одного микроба может быть множество различных антигенов, поэтому обычно вырабатывается целая серия антител, каждое из которых при этом направлено на определенный антиген. Антитела (иммуноглобулины, Ig) – это молекулы белков, способные прилипать к определенной структуре микроорганизма, вызывая его разрушение или скорейшее выведение из организма. Теоретически возможно формирование антител против любого химического вещества, имеющего достаточно большую молекулярную массу. Существует несколько типов иммуноглобулинов, каждый из которых выполняет специфическую функцию. Иммуноглобулины типа А (IgA) синтезируются клетками иммунной системы и выводятся на поверхность кожи и слизистых оболочек. В больших количествах IgA содержатся во всех физиологических жидкостях (слюна, молоко, моча). Иммуноглобулины типа А обеспечивают местный иммунитет, препятствуя проникновению микробов через покровы тела и слизистые оболочки.
Для характеристики микроорганизмов выделяют родовую, видовую, групповую и типовую специфичность антигенов. Наиболее точная дифференциация осуществляется с использованием моноклональных антител (МКА), распознающих только одну антигенную детерминанту.
Обладая сложным химическим строением, бактериальная клетка представляет целый комплекс антигенов. Антигенными свойствами обладают жгутики, капсула, клеточная стенка, цитоплазматическая мембрана, рибосомы и другие компоненты цитоплазмы, токсины, ферменты. Основными видами бактериальных антигенов являются:
- соматические или О- антигены (у грамотрицательных бактерий специфичность определяется дезоксисахарами полисахаридов ЛПС);
- жгутиковые или Н- антигены (белковые);
- поверхностные или капсульные К- антигены.
Выделяют протективные антигены, обеспечивающие защиту (протекцию) против соответствующих инфекций, что используется для создания вакцин.
Любой микроорганизм (бактерии, грибы, вирусы) представляет собой комплекс антигенов.
По специфичности микробные антигены делятся на:
· перекрестно-реагирующие (гетероантигены) - это антигены общие с антигенами тканей и органов человека. Они имеются у многих микроорганизмов и рассматриваются как важный фактор вирулентности и пусковой механизм развития аутоиммунных процессов;
· группоспецифические - общие у микроорганизмов одного рода или семейства;
· видоспецифические - общие у разных штаммов одного вида микроорганизмов;
· вариантспецифические (типоспецифические) - встречаются у отдельных штаммов внутри вида микроорганизмов. По наличию тех или иных вариантспецифических антигенов микроорганизмы внутри вида делят на варианты по антигенному строению - серовары.
По локализации антигены бактерий делятся на:
· целлюлярные (связанные с клеткой),
· экстрацеллюлярные (не связанные с клеткой).
Среди целлюлярных антигенов основными являются: соматический - О-антиген (глюцидо-липоидо-полипепдидный комплекс), жгутиковый - Н-антиген (белок), поверхностные - капсульные - К-антиген, Vi-антиген. Экстрацеллюлярные антигены - это продукты, секретируемые бактериями во внешнюю среду, в том числе антигены экзотоксинов, ферментов агрессии и защиты, и другие.
Антигены вирусов
В структуре вирусной частицы различают несколько групп антигенов:
· ядерные (или коровые)
· капсидные (или оболочечные)
На поверхности некоторых вирусных частиц встречаются особые V-антигены- гемагглютинин и фермент нейраминидаза.
Антигены вирусов различаются по происхождению. Часть из них – вирусоспецифические. Информация об их строении картирована в нуклеиновой кислоте вируса. Другие антигены вирусов являются компонентами клетки хозяина (углеводы, липиды), они захватываются во внешнюю оболочку вируса при его рождении путем почкования.
Антигенный состав вириона зависит от строения самой вирусной частицы. Антигенная специфичность простоорганизованныхвирусов связана с рибо- и дезоксирибонуклеопротеинами. Эти вещества хорошо растворяются в воде и поэтому обозначаются как S-антигены ( от лат. Solution- раствор). У сложноорганизованных вирусов часть антигена связана с нуклеокапсидом, а другая – локализуется во внешней оболочке – суперкапсиде. Антигены многих вирусов отличаются высокой степенью изменчивости. Это связано с постоянным мутационным процессом, который претерпевает генетический аппарат вирусной частицы. Примером могут служить вирус гриппа, вирысы иммунодефицитов человека.
14. Антигены гистосовместимости. На цитоплазматических мембранах практически всех клеток макроорганизма обнаруживаются антигены гистосовместимости. Большая часть из них относится к системе главного комплекса гистосовместимости, или МНС (аббр. от англ. Major histocompatibility complex).
По химической природе антигены гистосовместимости представляют собой гликопротеиды, прочно связанные с цитоплазматической мембраной клеток. Их отдельные фрагменты имеют структурную гомологию с молекулами иммуноглобулинов и поэтому относятся к единому суперсемейству.
Различают два основных класса молекул МНС. Условно принято, что МНС I класса индуцирует преимущественно клеточный иммунный ответ, а МНС II класса – гуморальный.
МНС I класса состоит из двух нековалентно связанных полипептидных цепей с разной молекулярной массой: тяжелой альфа-цепи и легкой бета-цепи. Альфа-цепь имеет внеклеточный участок с доменным строением (альфа1,альфа2,альфа3-домены), трансмембранный и цитоплазматический.
Для МНС I класса характерна высокая скорость биосинтеза – процесс завершается за 6 часов. Этот комплекс экспрессируется на поверхности практически всех клеток, кроме эритроцитов и клеток ворсинчатого трофобласта. Плотность МНС I класса достигает 7000 молекул на клетку, и они покрывают около 1% ее поверхности.
У человека МНС обозначили как HLA(аббр. от англ. Human Leukocyte Antigen), так как он ассоциирован с лейкоцитами.
В настоящее время у человека различают более 200 различных вариантов HLA I класса. Они кодируются генами, картированными в трех основных сублокусах 6-й хромосомы и наследуются и проявляются независимо: HLA-A, HLA-B, HLA-C. Локус А объединяет более 60 вариантов, В-130, а С- около 40.
Совершенствование или создание новых вакцин требует знаний структурных и функциональных особенностей вирусных антигенов, различаемых иммунной системой организма.
Вирусными антигенами называются продукты вирусспецифического синтеза, несущие признаки чужеродной генетической информации и вызывающие иммунный ответ. К ним относятся структурные и неструктурные вирусные белки.
Защита от вирусной инфекции зависит от выраженности иммунного ответа на антигены, расположенные на поверхности вирионов или инфицированных клеток. Иммунный ответ на неструктурные вирусные антигены играет меньшую роль в защите от инфекции.
Однако у герпесвирусов, например, клеточный иммунный ответ индуцируется множеством вирусспецифических белков, не входящих в структуру вирионов. Белки герпесвирусов эксперссируются каскадно и большинство нестук-турных белков синтезируется на ранней стадии репликации вируса. После про-цессинга они презентируются МНС классом I (главным комплексом гистосов-местимости, класс I) на плазматической мембране инфицированных клеток и распознаются специфическими цитотоксическими Т-клетками.
Поэтому инфицированные клетки могут различаться эффекторными цитотоксическими Т-лимфоцитами до завершения цикла вирусной репликации.
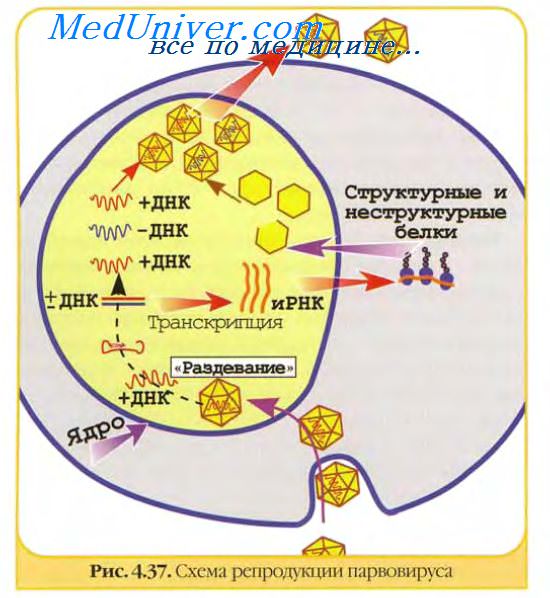
Каждый вирус представляет собой сложную смесь антигенов, определяемую в первую очередь структурными белками. Являясь сложными корпускулярными антигенами вирусы обычно вызывают выраженный иммунный ответ и большая часть их белков способна вызывать синтез специфических антител. Вирусные белки неравнозначны по своей антигенной активности. Наиболее явные и доступные мишени для иммунного ответа — белки, расположенные на поверхности вирусных частиц. Это прежде всего относится к вирусным гликопротеинам, расположенным на поверхности вирусных частиц, и экспрессированным на поверхности зараженных клеток.
Гликопротеины поверхности оболочечных вирусов и капсидные белки безоболочечных вирусов являются главными протективными антигенами.
Под специфичностью вирусного антигена подразумевают его способность избирательно реагировать с антителами или сенсибилизированными лимфоцитами, являющимися ответом на введение данного антигена. Участок антигена, который узнается специфическим лимфоцитом, и с которым впоследствии взаимодействует специфическое антитело, называется антигенной детерминантой.
Иммунологическая специфичность определяется не всей молекулой антигена, а лишь входящими в ее состав антигенными детерминантами (эпитопами). Участки вирусного белка, индуцирующие образование антител и специфически связывающиеся с ними, принято называть антигенными участками (доменами). Антитела соответствующей специфичности образуются к каждой антигенной детерминанте. Антитела к определенной детерминанте реагируют только с ней или с другой очень сходной структурой. Специфичность антигена определяется совокупностью детерминант, а его валентность — количеством однородных антигенных детерминант. Антигенность детерминант зависит от их пространственной структуры и размера молекулы антигена.
Антигенные детерминанты состоят обычно из 10—20 аминокислотных остатков и содержат гидрофильные группы. Наиболее гидрофильными аминокислотами являются лизин, аргинин, аспарагиновая кислота и глютаминовая кислота. Считается, что те участки молекулы белка, в которых их содержание относительно велико, предпочитают водное окружение и поэтому располагаются на поверхности. Различают линейные (непрерывные) и конформационные (прерывистые) детерминанты. Антитела образуются преимущественно к конформационным детерминантам, расположенным, как правило, на поверхности вирионов, и зависят от третичной структуры молекулы антигена.
Антигенная и иммуногенная активность вирусов определяется, главным образом, конформационными эпитопами. Разные антитела различают специфические антигенные участки вирусных антигенов. Например, прикрепительный гликопротеин (HN) вируса парагриппа имеет, по крайней мере, 6 антигенных сайтов, три из которых различаются нейтрализующими антителами.
Денатурация белков приводит к потере некоторых конформационных детерминант, обнажая ранее экранированные детерминанты. В результате денатурации белки частично или полностью изменяют антигенную специфичность, что может отразиться на иммунном ответе.
Вирионные белки разных вирусов различаются типоспецифичностью и вариабельностью. Одни из них обладают высокой вариабельностью, другие характеризуются консервативностью. Группоспецифические антигены являются высококонсервтивными, находятся обычно внутри вирионов и могут быть сходными у нескольких представителей рода данного семейства вирусов. Например, субвирусные частицы 12S вируса ящура содержат высококонсервативный белок, который выявляется моноклональными антителами одной специфичности у шести из семи известных типов вируса. Однако иммунизация ими не сопровождалась образованием ВН-антител.
Типоспецифические антигены связаны с вариабельными областями белков, обычно расположенными в наружных частях вирионов, и обладают узкой специфичностью, присущей одной группе вирусов.
Читайте также:
|
1. Структура вирусного генома
2. Генетические признаки вирусов
3. Изменчивость вирусов и селекция
4. Генная инженерия
Генетика наука о наследственности и изменчивости живых организмов.
Наследственные признаки очень стойкие и передаются, из поколения в поколение от родителей к потомкам через материальные носители наследственности – нуклеиновые кислоты.
Но в тоже время организмы в мире не изолированы, а взаимосвязаны с внешней средой и другими организмами. И под влиянием этих факторов происходят изменения отдельных признаков в организмах как на генетическом уровне (наследственная изменчивость), так и не связанные с изменением генетического аппарата (не наследственная изменчивость).
Наследственность и изменчивость тесно связаны между собой и являются противоположными сторонами единого процесса эволюции живых существ. Следует отметить, что вирусы в значительной мере способствовали развитию генетики как науки, так как они являются удобной моделью для изучения состава, репликации, функций нуклеиновых кислот, явлений изменчивости, что связано с простым строением и быстрым размножением (репродукция).
В развитии биологии в том числе и вирусологии и генетики важную роль сыграли такие открытия:
1. Расшифровка структуры молекулы ДНК
2. Расшифровка синтеза белка
3. Расшифровка генетического кода
Эти открытия сравнивают с открытием атомной энергии и ХХ век называют веком биологии в связи с тем, что были раскрыты интимные стороны жизни – тончайшее строение и функции нуклеиновых кислот.
1 Структура вирусного генома
Долгое время считалось, что носителем наследственной информации является белок. Но в 1944 г. Эвери доказал, что трансформации бактерий обусловлены нуклеиновой кислотой. В вирусологии значение нуклеиновой кислоты как носителя информации доказали Хергии и Чейз в 1952 году Они заметили что заражение бактерий может вызвать фаг, лишенный белковой оболочки. .Функцию вирусного генома выполняет или ДНК или РНК. ДНК или РНК могут быть одно или двухцепочные
Геном вирусов может быть представлен 10 вариантами:
1. Одноцепочной нефрагментированной РНК + цепью т.е. выполняет роль информационной РНК (пикорна- и тогавирусы, ретровирусы)
2. Одноцепочная нефрагментированная РНК – цепь ( рабдо-, парамиксовирусы)
3. Одноцепочная фрагментированная РНК – цепь (ортомиксовирусы)
4. Одноцепочная фрагментированная кольцевая РНК (бунъя вирус)
5. Двухцепочная фрагментированная РНК (реовирусы)
6.Одноцепочная ДНК нефрагментированная (парвовирусы )
7. Одноцепочная ДНК циркулярная (цирковирусы)
8.Двухцепочная ДНК нефрагментированная (герпес-, аденовирусы)
9.Двухцепочная ДНК кольцевая (папиломавирусы)
Соединяясь между собой в определенной последовательности, триплеты образуют генетический код.
Но количество возможных вариантов кодонов (64) превышает количество аминокислот (21)Это значит, что на каждую аминокислоту приходится 2-3 кодона для страховки, исключение - триптофан и метионин они кодируется одним триплетом.
Нуклеиновая кислота вирусов состоит из участков отличающихся друг от друга определенной последовательностью и количеством триплетов – эти участки называют генами и каждый ген ответственен за синтез одного определенного белка.
Число генов в нуклеиновой кислоте у разных вирусов варьирует в широких пределах. Например, вирус ящура и гриппа имеют по 3-5 генов у вируса полиоэмилита 10, а у вируса ньюкаслской болезни -37, вируса оспы-400 генов, а у человека-10 млн.
2 Генетические признаки вирусов.
Генетические признаки это любые наследственно-передаваемые свойства, которым можно дать качественную и количественную оценку, и которые проявляются в определенных условиях среды. Вирусы обладают определенными генетическими признаками, совокупность которых составляет генотип вируса, а совокупность проявленных генетических признаков составляет фенотип вируса.
Изучение генетических признаков вирусов имеет большое практическое значение, как в теоретическом, так и в практическом отношении. По этим признакам вирусы классифицируются, отбираются необходимые штаммы. Эти признаки или свойства используют в диагностической и лечебно-профилактической работе.
Но генетические признаки вирусов изучены недостаточно полно т. к. не все признаки, заложенные в генотипе, могут проявиться в конкретных условиях. И еще это связано с неоднородностью генотипов вирусных частиц в одной популяции.
Генетические признаки вирусов условно разделяют на: групповые, видовые и штаммовые.
Групповые признаки – это тип нуклеиновой кислоты, тип симметрии, размер и морфология, тип капсидной оболочки, количество капсомеров, наличие суперкапсидной оболочки, устойчивость к жирорастворителям или наличие липидов, антигенная специфика
Видовые признаки – патогенность для того или иного вида животного, тропизм, устойчивость к кислотности среды (рН), способность к гемагглютинации, характер ЦПД.
Штаммовые (внутривидовые) признаки - вирулентность для животных и куриных эмбрионов, терморезистентность, гемагглютинирующая активность, антигенная активность, характер бляшек, устойчивость к различным химическим и физическим факторам.
Каждый генетический признак вируса принято обозначать латинскими буквами:
Температура размножения rct - 40°
Инфекционная доза - ИД 50
Способность размножаться в культуре клеток, вызывая ЦПД – ТС
Устойчивость к рН среды - РН
3 Изменчивость вирусов и селекция.
В процессе репродукции вирусов в потомстве могут появляться вирионы, отличающиеся по своим свойствам от исходных родительских. Изменения могут касаться величины, формы патогенности, антигенной структуры, тропизма. Вновь возникшее свойства могут быть наследуемые т.е. связанные с изменениями в геноме вируса (генетические) или не наследуемые т.е. не связанные с изменениями в геноме вируса (фенотипические) .
Фенотепическая форма изменчивости или негенетическаясвязана с особенностями клетки хозяина, в которой происходит репродукция. У вирусов позвоночныхизменения связаны прежде всего со строением суперкапсида, который формируется на оболочках клетки . Поэтому при образовании оболочки включаются клеточные липиды, белки , углеводы. Так в оболочку вируса гриппа , культивированного на куриных эмбрионах , встраиваются белки алантоиса.
К фенотипическим формам изменчивости относят и негенетические взаимодействия вирусов между собой, это:
Фенотипическое смешивание происходит при заражении клетки двумя разными вирусами, при этом потомство преобретает признаки обоих вирусов. Например при заражении вирусом гриппа и вирусом нъюкаслской болезни . При этом в потомстве выявляются вирионы содержащие антигены и вируса гриппа и вируса нъюкаслской болезни.
Негенетическая реактивация (восстановление активности). при этом инактивированный вирус у которого разрушены белки -ферменты , необходимые для репродукции способен репродуцироватся за счет фермента у другого вируса даже у которого поврежен геном.
Комплементация это когда белки, кодируемые геномом одного вируса, способствуют репродукции другого вируса. Например предоставление фермента вирусу у которого его нет. Таким образом , комплементация приводит к формированию полноценного вириона, но она происходит только между близкородственными вирусами. комплементация может быть односторонняя , когда один вирус обеспечивает другого необходимыми продуктами для репродукции. Двусторонняя, когда каждый из вирусов не способен к самостоятельной репродукции.
К наследственно закрепленным формам изменчивости относят мутации и генетические взаимодействия вирусов. Причины сравнительно высокой способности вирусов к этому виду изменчивости , является огромная численность популяции вирусов, высокая скорость репродукции, слабая защищенность нуклеопротеида от внешних воздействий.
Мутация – это изменение последовательности нуклеотидов в определенном участке генома вируса.
В основе мутаций лежат следующие процессы:
1) Инверсия – изменение последовательности расположения одного или нескольких нуклеотидов (КОТ – ТОК) . Аналогично тому как меняется смысл вновь полученного слова, так и меняется состав гена, а значит при синтезе получится другой белок и другие свойства у вируса.
2) Замена одной или нескольких пар нуклеотидов другими( КОТ-КОМ).
3) Вставки – в цепь встраивается один или несколько нуклеотидов (ОКО-ОКНО).
4) Делеция – выпадение из цепи одного или нескольких нуклеотидов (ОКНО-ОКО)
5) Дупликация- дублирование одного или нескольких нуклеотидов
По обратимости необратимые при которых изменяется фенотип вируса и такие мутанты бястро вытесняют другиа популяции вируса.
Обратимые мутации при котрых происходит обратная мутация в месте первичной
По протяженности мутации могут быть точечными, захватывать лишь один триплет. Такие мутации могут не проявляться за счет того, что одна аминокислота кодируется– несколькими кодонами. Могут быть аберрационными , которые захватывают значительный участок гена. Такие мутации проявляются всегда.
По природе мутации бывают спонтанные и индуцированные.
Спонтанные – самопроизвольные, возникают в природе при воздействии на геном вируса различных естественных мутагенных факторов или ошибок действия ферментов ДНК-полимеразы или РНК-полимеразы
Одной из важных причин, приводящих к изменению вирусов в естественных условиях ,является коллективный иммунитет, который препятствует дальнейшему размножению вируса, вызвавшего инфекцию – (спад эпизоотии). В иммунном организме могут репродуцироваться антигенные варианты этого вируса, которые не обезвреживаются специфическими антителами. Следовательно, в процессе эпизоотии выживают вирионы с измененной антигенной структурой, которые в последствии после селекции образуют новую популяцию вируса, способную инфицировать иммунный организм.
Хорошо известна естественная изменчивость вируса гриппа, который проявляется появлением различных антигенных вариантов вируса. Способствующим фактором является фрагментированная РНК. В результате мутации и рекомбинации между вирусами гриппа человека и животных, образуются новые варианты вируса.
Вирус ящура имеет 7 типов, а внутри десятки вариантов и в ходе эпидемии происходит смена типов и вариантов, что затрудняет специфическую профилактику болезни.
Помимо антигенной изменчивости может наблюдаться изменчивость патогенных свойств – повышение или понижение вирулентности. Например вирус ньюкаслской болезни сначала вызывал смертельное заболевание птицы. В настоящее время регистрируют легкое течение данной болезни. такой вирус называется природно-ослабленный штамм и используется для приготовления вакцины. К сожалению, бывают и противоположные факты: усиление вирулентности вируса в природных условиях (так произошло с вирусом бешенства и вируса миксоматоза кроликов).
Индуцированные (искусственные)мутации – возникают в результате направленных воздействий экспериментатора на вирус различными физическкими и химическими мутагенами а также при адаптации вируса к необычной биосистеме.Такое воздействие на вирус вызывают мутаций в десятки и сотни раз эффективнее, чем природные факторы. Действие мутагенов имеет определённую направленность, что позволяет заранее предвидеть, куда действует мутаген и какие последствия вызовет.
1) Физические мутагены: повышенная температура способствует удалению пуринов из ДНК и замена другими; УФО – поглощается нуклеиновой кислотой, изменяет структуру пиримидинов
2).Химические мутагены могут действовать на нуклеиновую кислоту во время её репликации (аналоги пуриновых и пиримидиновых оснований) или вступать в реакцию с покоящейся молекулой нуклеиновой кислотой, но требующие для выявления (формирования) мутаций, последующей её репликации (азотистая к-та, гидроксиломин) и т.д.
Пастер 1822-1895 – ослабил вирус бешенства, пассируя через организм кролика.
Рекомбинация – это обмен генетическим материалом между двумя близкими, но отличающимися по наследственным свойствам вирусами.
Рекомбинации могут быть:
межгенные – обмен полными генами,1 внутригенная – обмен участками генов. Образующися рекомбинантный вирус преобретает свойства обоих вирусов. Рекомбинанты вирусов получаются только при скрещивании близких по свой ствам вирусов родственных.
Рекомбинации между одноцепочными НК и между двухцепочными НК
В экспериментальных условиях гибридные (рекомбинантные) формы можно получить при совместном введении в клетку:
1) двух жизнеспособных вирусов ;
2) живого и инактивированного вируса
3) живого вируса и вирусной нуклеиновой кислоты , выделенной из другого штамма;
4) одновременно двух нуклеиновых кислот от разных вирусов.
Таким образом, получают гибриды с признаками не встречающиеся в природе. Так в результате гибридизации инактивированного УФ-лучами вируса гриппа типа А, образующего бляшки, с инфекционным вирус гриппа не обладающего такой способностью получили бляшкообразующий инфекционный вирус. Открыты гибридизации между вирусами гриппа животных и человека, ящура и реовирусов.
К генетическим взаимодействиям вирусов относят также
множественную реактивацию( восстановление активности), которая наблюдается при заражении клетки несколькими вирионами с поврежденным геном. При этом функцию повркжденного гена выполняет вирус, у которого этот ген не поврежден . Таким образом вирионы дополняют друг друга путем генных рекомбинаций и в результате репродуцируется неповрежденный вирус.
Кросс-реактивация-Сходна с предыдущей но отличается тем что один вирус используют в неизменным а другой инактивируют при этом наблюдается восстановление активности инактивированного генома геномом неповрежденного вируса
Пересортировка генов наблюдается среди вирусов с фрагментированным геномом. При этом образуются группировки генов которые позволяют более благоприятно жить вирусу в данной биосистеме.
Гетерозиготность. феномен , наблюдающийся в случае репродукции в клетке вирионов отличных по генетическим признакам. При этом потомство может обладать частично свойствами одного, частично другого родителя и частично смешаными свойствами ( вирус гриппа и вирус болезни Нъюкасла)
Транскапсидация наблюдается при репродукции в клетке нескольких неродственных вирусов. При этом вирусный геном неспособный сентезировать белки капсида прекрепляется в неизменном виде к геному другого вируса и "переезжает" сним в другую клетку.
Популяции вирусов в результате генетических взаимодействий: рекомбинаций, пассажей или воздействия мутагенов и тд. генетически неодинаковы. В общей массе мутированных, есть вирионы, которые не изменились или реверсировали (восстановили свои свойства). В результате дальнейшей репродукции ( за счет них) популяция вирусов может восстанавливать свои первичные свойства.
Поэтому проводят селекцию вирусов, используя различные методы отбора вирионов и создание однородной популяций .
1. Выделение культуры вируса из отдельных бляшек на ХАО куриного эмбриона или на культуре клеток . Так как считается, что 1 бляшка это потомство 1 вириона.
2. Методом предельных разведений . Вирус разводят до 1 вириона в определенном объеме культуральной жидкости и этой дозой заражается культура клеток или куриный эмбрион. В результате получается популяции из одного вириона.
3. Методом избирательной адсорбции и элюции. Вирусную суспензиюсмешивабт с сорбентом ,на который прилипают только измененные вирионы. Затем сорбент с вирионами отмывают , а вирионы снимают с сорбента .
4. Методом пассажей в измененных условиях культивирования. вирусы продолжают культивировать на нечуствительной живой системе в течение 100-200 пассажей . за это время останутся только измененные вирионы , которые уже не смогут восстановить свои первоначальные патогенные свойства , даже если их ввести в бывшую чуствительную систему
4. Генная инженерия
Благодаря детальному изучению структуры ДНК, синтеза белка появилась отрасль науки – генная инженерия – цель которой создание новых генетических структур. В отношении вирусологии эта отрасль работает в направлении:
1. Создание новых вирусных геномов.
2. Получение вирусных протективных белков , на которые образуются защитные антитела в организме и получение интерферона.
Конечной целью первого направления является создание новых вакцин против вирусных болезней.
Если ввести такой рекомбинант в организм он будет вызывать образование защитных антител к двум вирусам. Таким образом получают рекомбинантные вакцины.
Для получения определённого вида белка например интерферрона или антигенных белков вируса поступают аналогично. Получают ген из природной ДНК вируса или клетки ,ответственный за синтез белка и встраивают в вектор (вирус, фаг, плазмида).
Вектор вводят в клетку в которой вектор размножается, давая необходимое количество геномов содержащих ген ответственный за синтез определенного белка.
Затем под различными воздействиями геном запускает синтез этого белка и он получается в необходимых количествах (интерферон, субъединичные вакцины ,инсулин и т.д.)
Таким образом в промышленных масштабах можно получать дешовые белковые препараты необходимые в любой отрасли (медицина, ветеринария, пищевая промышленность и тд).
Читайте также:
