
У паразитов анаэробное дыхание
Дыхание эктопаразитов подчиняется тем же законам, что и дыхание свободно живущих организмов. Полостные и тканевые паразиты находятся, очевидно, в таких же условиях дыхания, как и соседние ткани самого хозяина, т. е. кислород доставляется им кровью хозяина или (у Arthropoda Tracheata и у паразитов, живущих в дыхательных путях позвоночных) непосредственно органами дыхания хозяина. Совершенно особые условия дыхания окружают кишечных паразитов: они живут в почти бескислородной среде.
В течение долгого времени считалось, что кишечные .паразиты обладают чисто анаэробным типом обмена веществ, т. е. что источником энергии у них являются процессы распада (брожения) углеводов и жиров, протекающие без использования свободного кислорода. Так, Вейнланд (Weinland, 1901) давно уже показал, что у аскариды в тканях в большом количестве накапливается гликоген и источником энергии является его расщепление, происходящее с выделением углекислоты и валериановой кислоты. Однако в настоящее время можно считать доказанным (см., например, Брандт, 1951; Brandt, 1952), что у кишечных паразитов наряду с анаэробными процессами брожения (расщепления) имеют место и аэробные процессы, связанные с использованием свободного кислорода. Такого рода смешанный тип обмена носит название аноксибиоза.
Анаэробиоз благодаря отсутствию ресинтеза продуктов брожения характеризуется значительно меньшей энергетической эффективностью, чем оксибиоз. При аноксибиозе в зависимости от большей или меньшей степени преобладания анаэробных процессов над аэробными энергетическая эффективность может быть различной. Например, у аскариды в процессе обмена веществ освобождается примерно 9% той энергии, которая освободилась бы при полном окислении углеводов. Тем не менее по сравнению с анаэробными формами брожения у микроорганизмов (бактерий, дрожжей), анакси- биоз паразитических червей характеризуется все же относительно высокой энергетической эффективностью. Так, освобождение энергии при маслянокислом брожении равно всего 2,2%, а при спиртовом брожении — 4% (по сравнению с полным окислением).
В теле паразитов-аноксибионтов почти всегда имеются значительные запасы гликогена, который и является основным энергетическим источником.
Экспериментально доказано (Марков, 19396, 1943, 1958), что при переживании паразитов вне тела хозяина в растворах, лишенных питательных веществ (например, раствор Рингера), происходит трата гликогена. Чем выше температура и чем активнее движение паразита, тем быстрее потребляются запасы гликогена. Кроме гликогена в тканях паразитов-аноксибионтов обычно отлагаются значительные количества жира, который является конечным продуктом при брожении гликогена (Brandt und Weinland, 1924; Лутта, 1939; Марков, 1950, 1958).
В некоторых случаях наблюдаются интересные биологические приспособления внутренностных паразитов, облегчающие им получение атмосферного воздуха. Так, нами уже говорилось, что личинки кожного овода, располагаясь под кожей рогатого скота, устанавли- в 2ют связь с внешней средой при помощи прободающего.. кожу отверстия. Другой интересный пример дает представитель паразитических мух Crossocosmia (Ugimyia) sericariae ( 109). Яйца этой мухи откладываются ею на листья тутового дерева и поедаются б месте с пищей гусеницами тутового шелкопряда. Личинки мухи вылупляются из яиц в кишечнике гусеницы, прободают стенку кишки :: сначала внедряются в ганглии брюшной нервной цепочки хозяина. Вскоре они, однако, покидают ганглий и выходят в полость тела гусеницы, где располагаются так, чтобы их находящиеся на заднем конце тела дыхальца соприкасались с одним из дыхалец хозяина.
Подобное положение паразитов настолько постоянно, что о присутствии их внутри гусениц шелкопряда узнают по наличию около одного из дыхалец хозяина буроватого пятна, вызываемого паразитом. Очевидно, в этом тесном контакте со стигмой хозяина мы имеем дыхательное приспособление личинок Crossocosmia (Ugimyia).
Очень интересные приспособления для дыхания обнаруживают некоторые вши морских млекопитающих, а именно: ластоногих. Некоторые виды вшей ластоногих не имеют специальных морфологиче- ; еких а'даптаций и располагаются преимущественно на морде своего хозяина, т. е. на том месте тела, которое чаще всего выставляется хозяином из воды на воздух. Здесь, следовательно, мы имеем биологическое приспособление паразита, в смысле его локализации. У других вшей, относящихся к роду Antarctophthirius, на теле развиваются особые плоские чешуйки ( 110). При погружении хозяина в воду эти чешуйки прижимаются насекомыми к его телу, зажимая в пространстве между чешуйкой и телом маленький пузырек воздуха. Таким путем паразит получает на время погружения хозяина некоторый запас уносимого под воду воздуха. Подобное морфологическое приспособление позволяет Antarctophthirius жить на всей поверхности тела хозяина.
Некоторые веслоногие рачки, прячущие большую часть своего тела внутрь организма хозяина (таков Herpyllobius), имеют полостную жидкость розоватого цвета.
Интересующим нас в данном случае обстоятельством является го, что большей частью у свободно живущих и свободно дышащих воздухом родичей таких паразитов эритрокруорин отсутствует, т. е. приобретение его паразитами означает собою одну из адаптаций к.парази- ческому образу жизни. Очень хорошо обосновывает эту мысль Кемниц сравнением личинок желудочного овода с таковыми бычьего (Hypoderma lineatum). Личинки бычьего овода живут в подкожных желваках, сообщающихся с внешней средой, и дышат наружным воздухом; их трахейные клетки лишены красного пигмента, тогда как личинки желудочного овода отрезаны от свободного доступа воздуха и имеют эритрокруорин. Таким же образом свободно живущие нематоды не обнаруживают красной окраски, а у некоторых эидопаразитных круглых червей последняя имеется. По-видимому, эритрокруорин некоторых эндопа- < разитов Содействует им в поглощении кислорода из бедной этим газом окружающей среды.
Выделение излишней воды (осморегуляция), как и у всех других простейших, осуществляется при помощи сократительных вакуолей, которые имеют более сложное, чем у
Лейшмании являются внутриклеточными паразитами, поражающими систему мононукле- арных фагоцитов.
Клеточное дыхание — это окисление органических веществ в клетке, в результате которого синтезируются молекулы АТФ. Исходным сырьем (субстратом) обычно служат углеводы, реже жиры и еще реже белки. Наибольшее количество молекул АТФ дает окисление кислородом, меньшее – окисление другими веществами и переносом электронов.
Углеводы, или полисахариды, перед использованием в качестве субстрата клеточного дыхания распадаются до моносахаридов. Так у растений крахмал, а у животных гликоген гидролизуются до глюкозы.
Глюкоза является основным источником энергии почти для всех клеток живых организмов.
Первый этап окисления глюкозы — гликолиз. Он не требует кислорода и характерен как при анаэробном, так и аэробном дыхании.
Биологическое окисление
Клеточное дыхание включает в себя множество окислительно-восстановительных реакций, в которых происходит перемещение водорода и электронов от одних соединений (или атомов) к другим. При потери электрона каким-либо атомом происходит его окисление; при присоединении электрона — восстановление. Окисляемое вещество — это донор, а восстанавливаемое — акцептор водорода и электронов. Окислительно-восстановительные реакции, протекающие в живых организмах носят название биологического окисления, или клеточного дыхания.
Наиболее распространенным в природе конечным акцептором электронов является кислород, который восстанавливается. При аэробном дыхании в результате полного окисления органических веществ образуются углекислый газ и вода.
Биологическое окисление протекает по-этапно, в нем участвуют множество ферментов и соединения, переносящие электроны. При ступенчатом окислении электроны перемещаются по цепи переносчиков. На определенных этапах цепи происходит выделение порции энергии, достаточной для синтеза АТФ из АДФ и фосфорной кислоты.
Биологическое окисление весьма эффективно по-сравнению с различными двигателями. Около половины выделяющейся энергии в конечном итоге фиксируется в макроэргических связях АТФ. Другая часть энергии рассеивается в виде тепла. Поскольку процесс окисления ступенчатый, то тепловая энергия выделяется понемногу и не повреждает клетки. В то же время она служит для поддержания постоянной температуры тела.
Аэробное дыхание
Различные этапы клеточного дыхания у аэробных эукариот происходят
в матриксе митохондрий – цикл Кребса, или цикл трикарбоновых кислот,
на внутренней мембране митохондрий – окислительное фосфорилирование, или дыхательная цепь.
На каждом из этих этапов из АДФ синтезируется АТФ, больше всего на последнем. Кислород в качестве окислителя используется только на этапе окислительного фосфорилирования.
Суммарные реакции аэробного дыхания выглядит следующим образом.
Дыхательная цепь: 12H2 + 6O2 → 12H2O + 34АТФ
Таким образом биологическое окисление одной молекулы глюкозы дает 38 молекул АТФ. На самом деле нередко бывает меньше.
Анаэробное дыхание
Большинство анаэробов — это микроорганизмы. Однако к организмам, использующим анаэробное дыхание, относятся также дрожжи, ряд червей-паразитов. Способностью к анаэробному дыханию также обладают определенные ткани. Например, мышечные клетки, которые периодически могут испытывать недостаток кислорода.
При анаэробном дыхании в окислительных реакциях акцептор водорода НАД не передает водород в конечном итоге на кислород, которого в данном случае нет.
В качестве акцептора водорода может быть использована пировиноградная кислота, образующаяся при гликолизе.
У дрожжей пируват сбраживается до этанола (спиртовое брожение). При этом в процессе реакций образуется также углекислый газ и используется НАД:
CH3COCOOH (пируват) → CH3CHO (ацетальдегид) + CO2
Молочнокислое брожение происходит в животных клетках, испытывающих временный недостаток кислорода, и у ряда бактерий:
CH3COCOOH + НАД · H2 → CH3CHOHCOOH (молочная кислота) + НАД
Оба брожения не дают выхода АТФ. Энергию в данном случае дает только гликолиз, и составляет она всего две молекулы АТФ. Значительная часть энергии глюкозы так и не извлекается. Поэтому анаэробное дыхание считается малоэффективным.
Эта статья уделяет внимание различным видам червей, в частности плоским, круглым и кольчатым. Особое место будет выделено плоским червям. Будут рассмотрены их различные органы и их деятельность. Например, мы разберем, как осуществляется дыхание плоских червей, изучим строение выделительной и репродуктивной систем и т.д. А также будут рассмотрены некоторые их представители.
Разнообразие червей

К типу Черви относится группы многоклеточных животных, у которых вытянутое тело и отсутствует скелет. Места обитания - это, как правило, влажная почва, море и пресные водоемы. По размерам они могут варьироваться от таких, которые можно обнаружить лишь в микроскоп, до крупных форм, длинной в несколько метров. В соответствии с формой тела выделяют: Плоских, Круглых и Кольчатых червей. У всех типов имеется три слоя тела. Зародышевые слои – эктодерма, эндодерма и мезодерма дают начало развитию всем их тканям и органам.
Самые яркие и известные представители плоских червей: планария, печеночная двуустка, свиной и бычий цепни, эхинококки, шистосома и т.д. К известным кольчатым относятся: червь дождевой, малощетинковые черви, пиявки и мизостомиды. Круглые первичноротые представлены всем известными аскаридами, острицами, риштами, трихинеллами и т.д.
Несмотря на разнообразие существующих видов червяков, их типов, особенностей строения, способов репродукции, питания, мест обитания и т.д., существуют немалое количество сходств, характерных для них всех. Например, дыхание плоских червей, разделенное на аэробное и анаэробное, в зависимости от среды обитания, характерно и для других двух типов.
Плоские черви

Начнем с общей характеристики червей. Плоские черви – это беспозвоночные животные, относящихся к первичноротым. Данные существа относятся в таксономической иерархии к животным многоклеточного типа, имеющих вытянутую форму тела и отсутствие внутреннего скелета. Зоология типа Плоские черви представляет собой описание строения, процессов жизнедеятельности и физиологии этих существ. Они являются обитателями соленых и пресных водоемов, другие представители могут выживать в местах с высокой влажностью. Остальные классы занимаются паразитизмом, живя на разнообразных животных, включая и позвоночных и беспозвоночных. Сейчас описано приблизительно 25000 видов, на территории РФ обитает свыше трех тысяч видов.
Система органов плоских червей представлена некоторым количеством основных структурных компонентов, объединенных по общим функциональным признакам и по типу строения. К основным системам относятся: дыхательная, репродуктивная, выделительная, мышечная, нервная и покровная.

Некоторые представители плоских червей, например, планария – населяют пресные водоемы. Среди ресничных червей это наиболее известный. К паразитам относятся сосальщики, например, печеночные и кошачьи двуустки, шистосомы, а также ленточные (широкие лентецы, бычьи и свиные цепни, эхинококки).
Ранее к классу ресничных первичноротых приписывали ряды других таксономических элементов, характеризующихся червеобразными формами, отсутствием полостей тела и считающимися беспозвоночными.
Форма тела у любого типа, имеет билатерально-симметрическую форму, в которой выражены головной и хвостовой концы, слегка уплощенные оба конца, однако у крупных видов, уплощение сильно выраженно. Система органов плоских червей для дыхания и кровообращения – отсутствует. Полость в теле не развивается, однако это справедливо для всех представителей, кроме ленточных червей и сосальщиков на определенных жизненных циклах.
Строение покровов тела
Дыхание плоского червя осуществляется именно через поверхность тела организма, потому оно связанно со строением покровов организма. Снаружи тело покрывается одним слоем эпителия. Ресничные черви (турбеллярии) располагают эпителием, состоящим из клеток, что несут реснички. Плоские черви-паразиты сосальщики, а также представители моногенеев, цестодообразных и ленточных червей в течение основной части своей жизни не обладают ресничным эпителием. Клетки ресничного типа могут встречаться у личинок. Покровы тела этих трех типов представляются в виде тегументов, которые несут на себе микроворсинки или хитиновые крюки. Обладателей тегумента называют представителями группы Neodermata. Около 6/7 составной своего тела плоские червы способны восстанавливать путем регенерации.
Знакомство с мускулатурой

Мышечные ткани плоских червей представлены мускульным мешком, который залегает под эпителием. Он состоит из некоторого количества слоев клеток мышечного типа, которые не разделены на мышцы. Однако некоторая дифференциация наблюдаема в областях глотки и половой системы. Внешняя часть клеток мышечных слоев ориентируются поперек, а внутренние – вдоль задне-передней оси тела. Наружная мускулатура называется кольцевым слоем, а внутренняя – слоем мускулатуры продольного характера.
Способы дыхания
Сейчас мы постараемся разобрать вопрос о том, как осуществляется дыхание плоских червей? Детальное описание процессов дыхания описывается лишь поверхностно. Важно знать лишь то, что дыхание плоских червей осуществляется через полость всего тела. Из этого следует, что особые органы дыхания, свойственные многим животным, у них отсутствуют. Однако это касается паразитических форм червей и свободноживущих видов, а эндопаразиты, проживающие в среде с малым количеством кислорода, могут осуществлять дыхание анаэробным способом.
Дыхание плоских червей аэробного типа осуществляется путем диффузии – взаимопроникновении, например, газов, с целью выравнивания их по всему объему тела. Анаэробный гликолиз эндопаразитов представляет собой процесс самодостаточного типа, которому характерно наличие трех условий: прибытие глюкозы, наличие АТФ, практически в любом количестве и восстановление утерянного запаса НАД.
Ознакомление с глоткой и кишкой
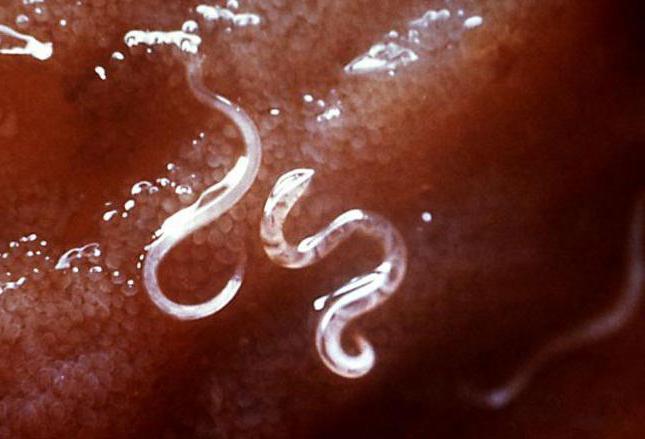
Всем группам плоских червей свойственно наличие глотки, которая ведет в кишку. Исключение составляют цестодообразные и ленточные червяки. Эта кишка открывается в паренхиму, предназначенную для пищеварения, слепо замыкается и связана с окружающим миром лишь ротовым отверстием. Некоторые крупные турбеллярии имеют в своем наличие анальные поры, однако, это исключение лишь для некоторых субъектов вида. Мелкие формы характеризуются прямым кишечником, а крупные (планария, сосальщик) могут иметь разветвленный. Глотка расположилась на поверхности брюха, часто ее можно обнаружить посреди или ближе к задней части тела. У некоторых групп червей глотка смещается вперед.
Особенности нервной системы и органов чувств
Характеризуя нервную систему плоских первичноротых, стоит отметить, что им характерно наличие нервных узлов, расположенных спереди тела, а также есть ганглии мозга и ответвляющиеся от них нервные столбцы, которые соединяются перемычками. К чувствительным органам относятся отдельные кожные реснички, которые являются отростками клеток нервного типа. Существуют свободноживущие виды, которые имеют особые, чувствительные к свету глазки пигментной природы. Такие органы служат примитивным приспособлением к ощущению равновесия и позволяют видеть, пусть и примитивно.
Система выделения

Репродукция
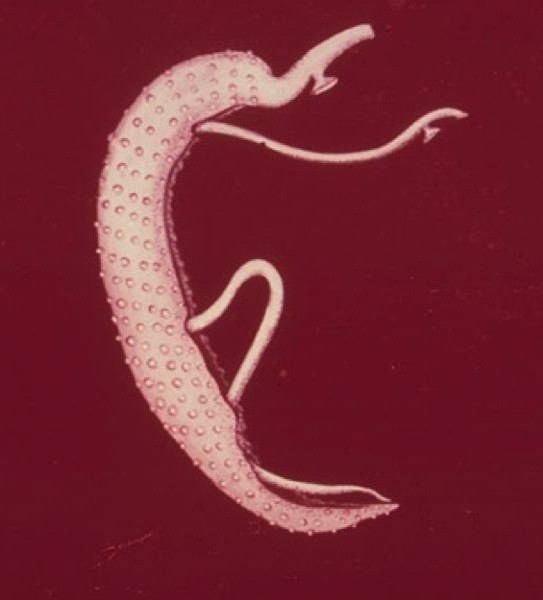
Среди плоских червей преобладают гермафродиты, лишь некоторые виды являются раздельнополыми, например, schistosomatidae. Половая система, как мужская, так и женская, может сильно отличатся между видами в плане формы строения семенников и яичника. Тоже касается и остальных составных половой системы. Определенные группы ресничных червей и все представители паразитов имеют яичник, разделенный на 2 части:
- Гермарий – является собственно яичником. Производит яйцеклетки, бедные на желток, но способные к развитию.
- Вителлярий – иногда называют желточником, он производит яйцеклетки абортивного типа, они богаты желтком.
Данные составные репродуктивно системы формируют сложные, или экзолецитальные, яйца. Общая оболочка может содержать в себе одну яйцеклетку или некоторое количество желточных шаров, выделяемых железами придаточного типа.
Заключение
Подытожив вышенаписанный текст, можно сделать несколько заключений, среди которых самыми значимыми являются: дыхание плоских червей осуществляется поверхностью всего тела, преимущественно плоские червяки относятся к хищникам, имеется мускульный мешок, покров тела представлен тегументом, большинство относятся к гермафродитам и лишь немногие из них раздельнополые.
Тип круглые черви эволюционно происходит от плоских червей - круглые черви также трехслойные, обладают несегментированным телом, билатеральной (двусторонней) симметрией, кожно-мускульным мешком. Возникновение круглых червей сопровождается рядом крупных ароморфозов, которые мы непременно с вами обсудим.
Представители типа: аскарида, острица, ришта, трихинелла.

- Первичная полость тела
У круглых червей впервые возникает полость тела! Отсюда важное наименование, применимое к типу круглые черви - первичнополостные.
Только представьте - у плоских червей пространства внутри тела не было, все заполняла клеточная масса - паренхима (мезенхима). У круглых червей появилось пространство между органами, заполненное межклеточной жидкостью.
Первичная полость тела, которая характерна для этого типа, по-другому называется - протоцеле, схизоцель, псевдоцель. Эта полость, как мы уже с вами убедились, заполнена межклеточной жидкостью - именно поэтому тело круглых червей на поперечно срезе всегда округлое (в отличие от тела плоских червей, которое "плоское").
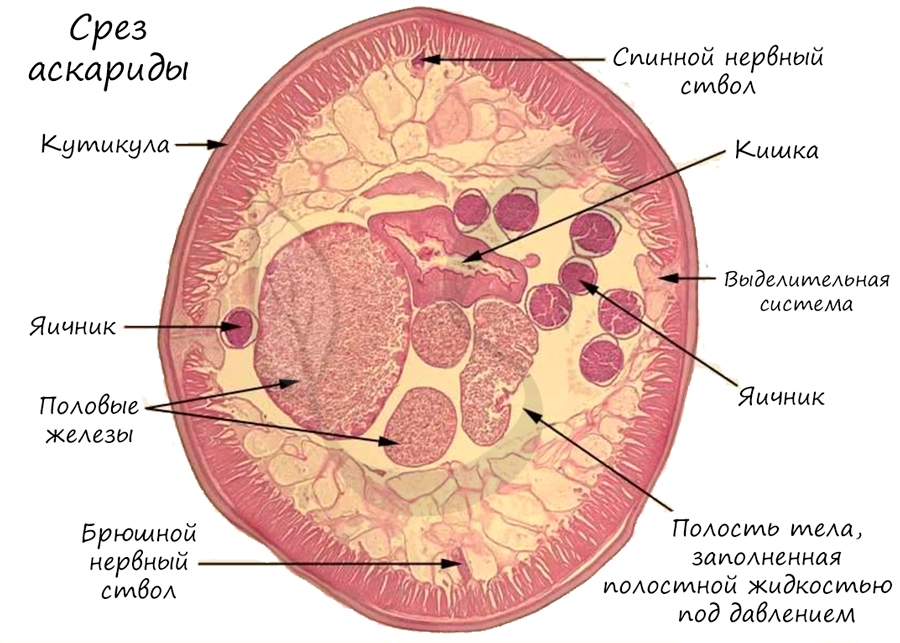
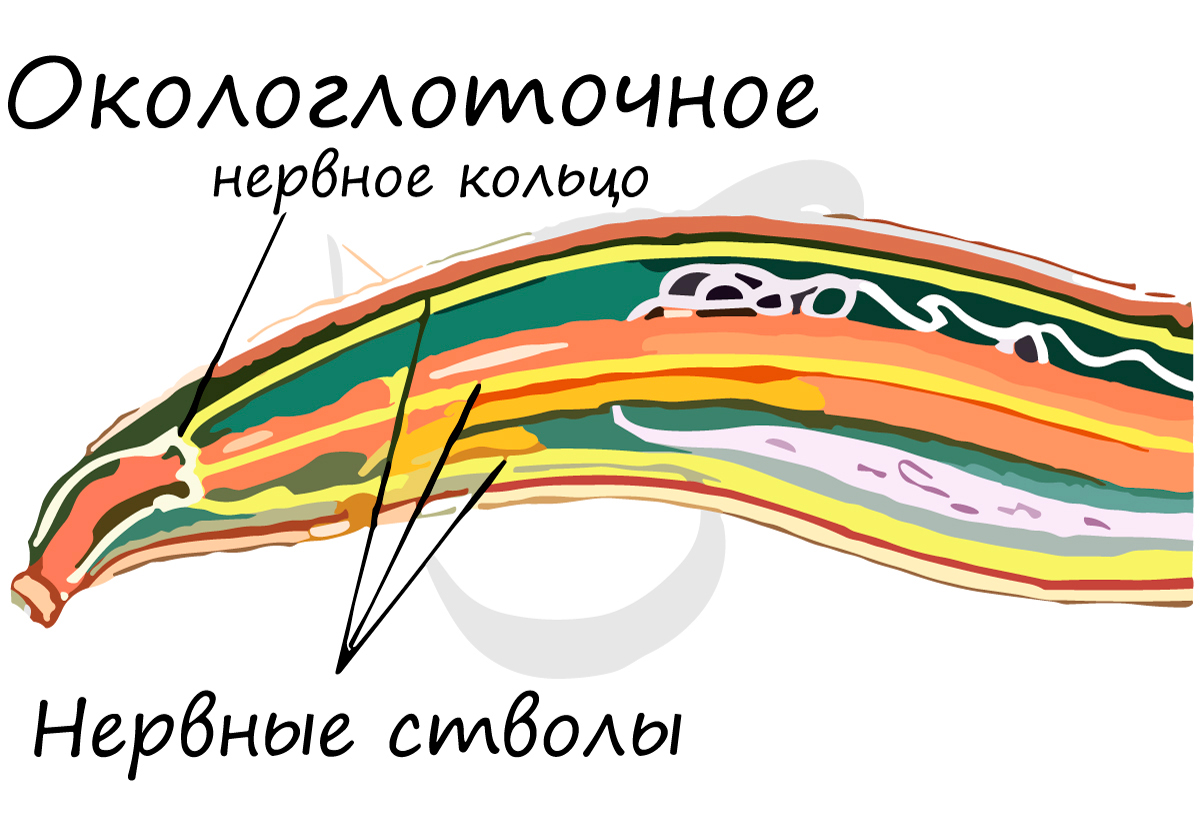
У круглых червей нервная система более развита по сравнению с предшествующими, изученными нами типами. Головные нервные узлы (ганглии) соединены друг с другом множеством перемычек, образуя окологлоточное нервное кольцо. От него отходят спинной и брюшной нервные стволы, соединенные поперечными перемычками - комиссурами.
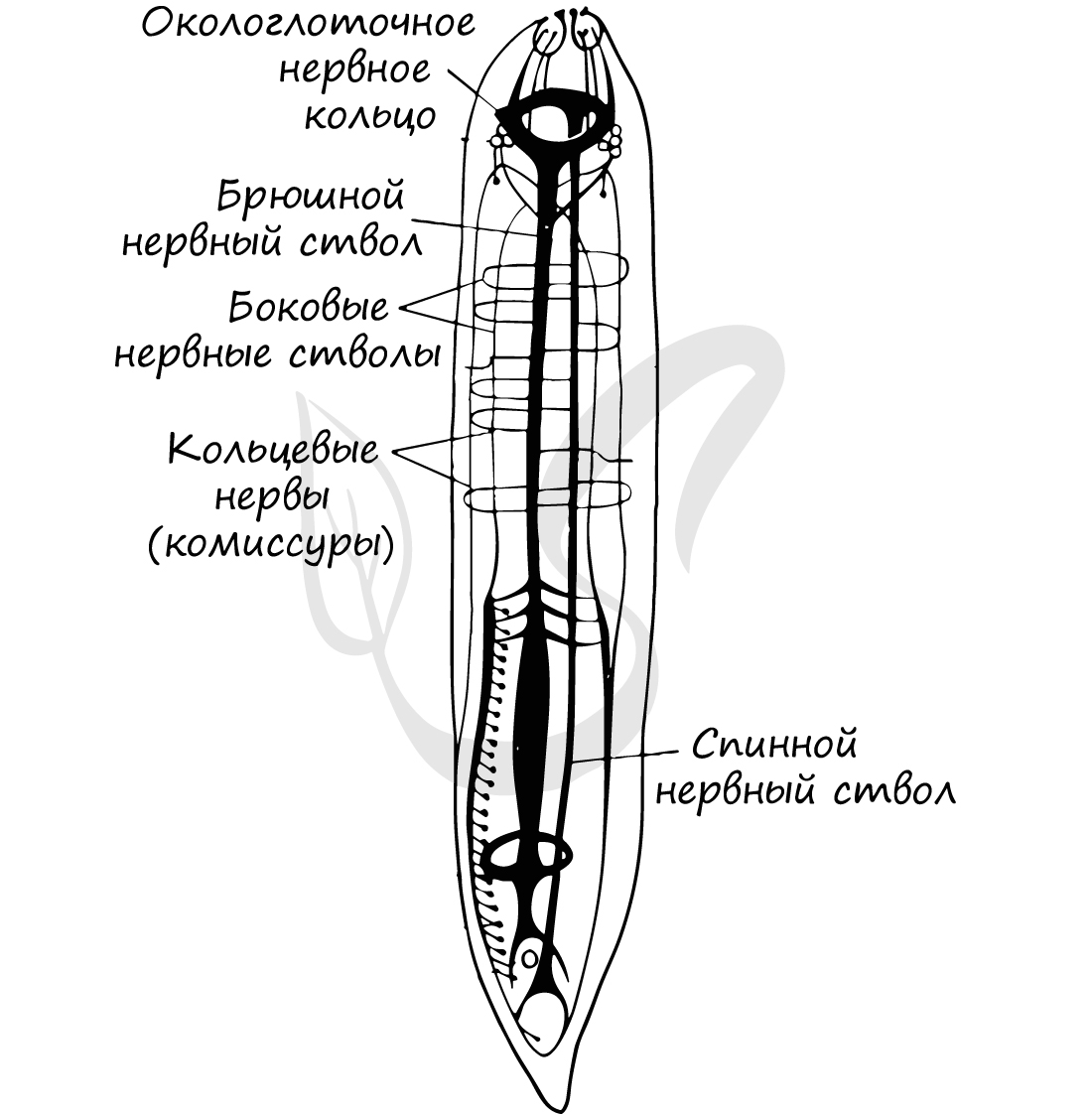
Пищеварительная система претерпевает резкие изменения: появляется задний отдел кишечника и впервые - анальное отверстие. Как вы помните, кишечнополостные и плоские черви из-за его отсутствия были вынуждены выделять непереваренные остатки пищи через рот - это эволюционно осталось позади. Теперь у пищеварительной системы появляется возможность функционировать непрерывно.
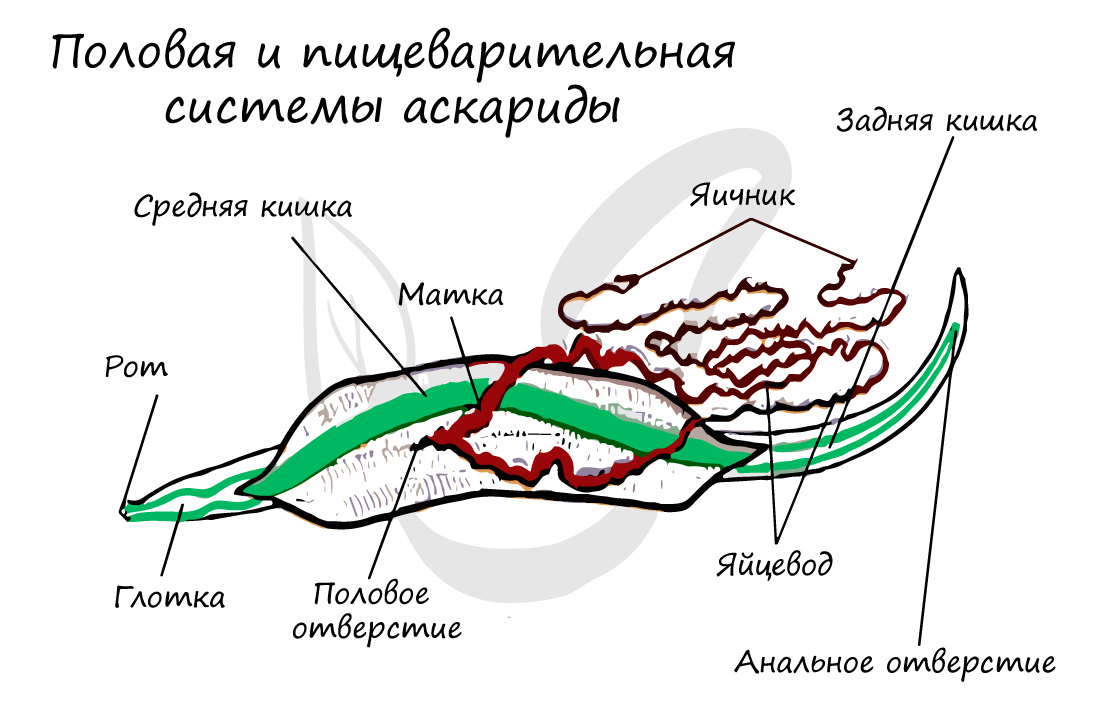
Продольная мускулатура дифференцируется (делится) на четыре мышечных тяжа: брюшной, спинной и два боковых. Благодаря такому строению становится возможным змеевидное движение.
Мужские половые органы находятся у мужских организмов, а женские - у женских. Это создает основу для комбинативного разнообразия потомства.
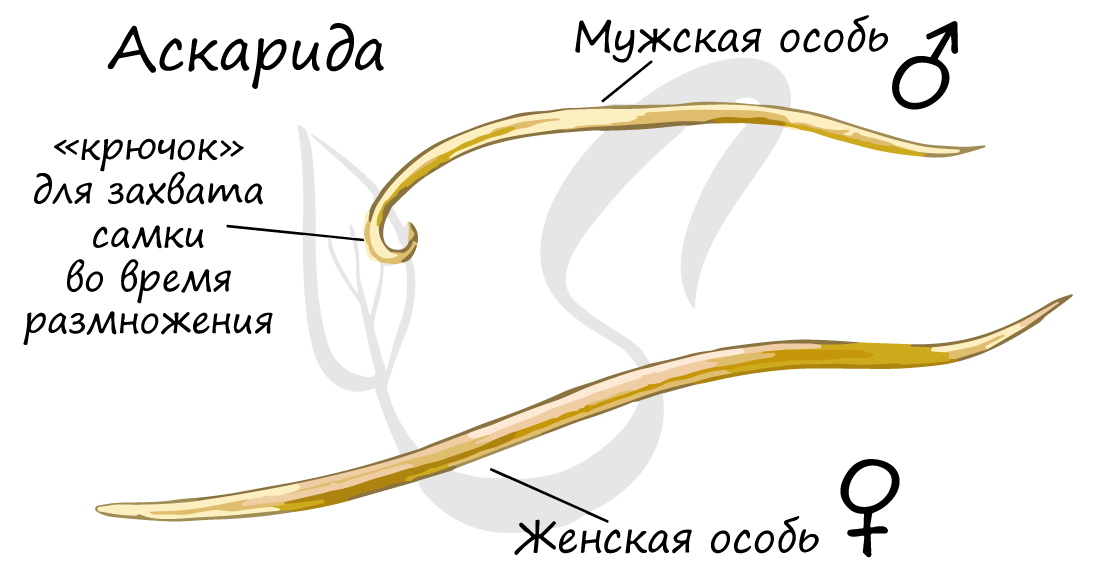
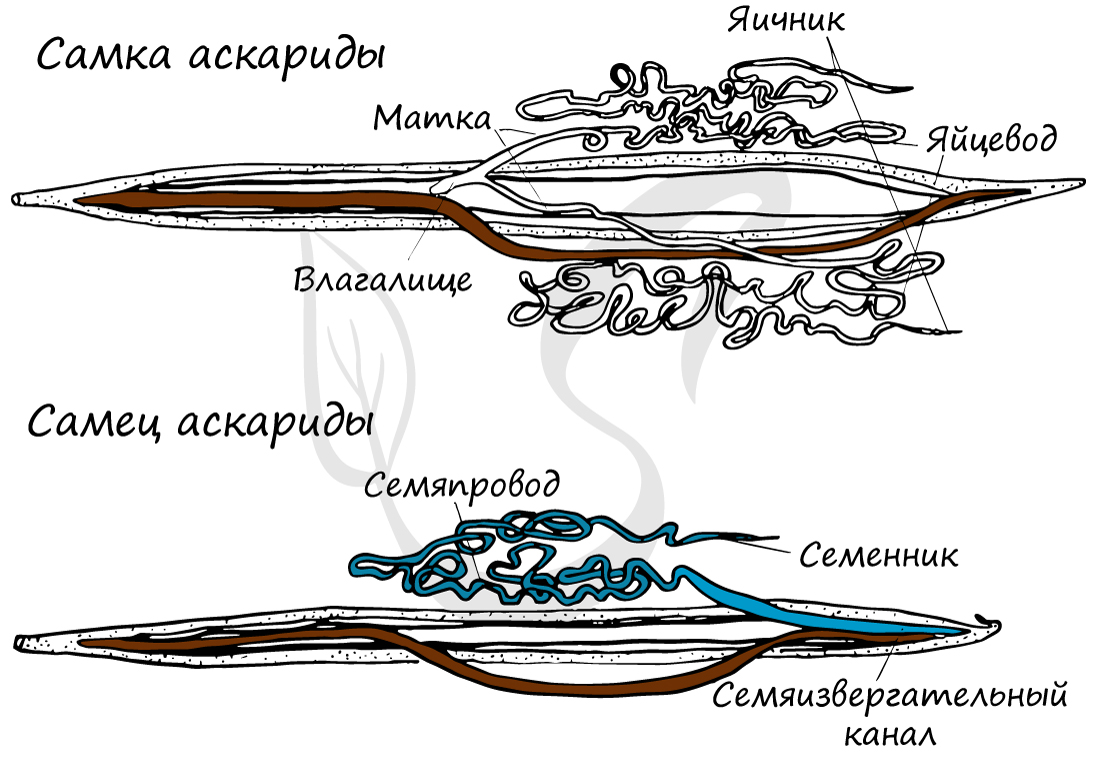
Обратите внимание (картинка ниже) - как хорошо у аскариды выражен половой диморфизм - это свойство имеется и у многих других представителей круглых червей. Половой диморфизм (от греч. di - две и morphe - формы) - внешние признаки различия между мужской и женской особью.
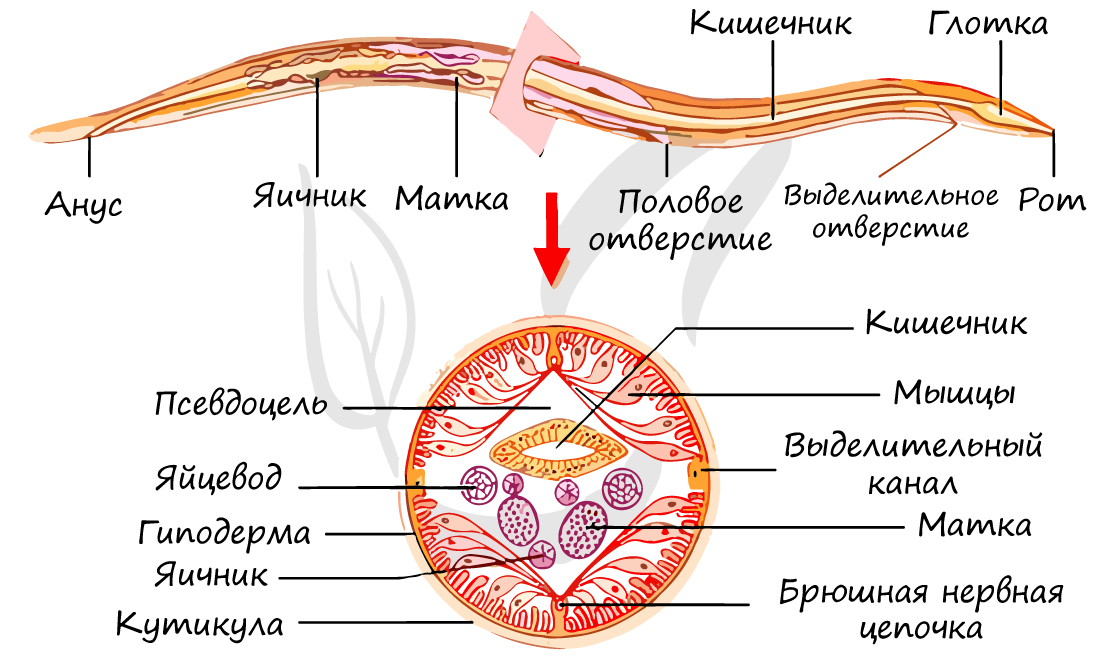
- Покровы тела и опорно-двигательная система
Покровные ткани кожно-мускульного мешка круглых червей представлены многослойной кутикулой и гиподермой. Кутикула (от лат. cuticula - кожица) - производное клеток однослойного покровного эпителия гиподермы. Представляет собой своеобразный "каркас" для мускулатуры. Важнейшая функция кутикулы - защита червя от переваривания ферментами кишечника, что дает возможность паразиту жить долгие годы в организме-хозяине.
Под кутикулой расположена гиподерма. Гиподерма имеет 4 протяженных утолщения, тянущихся вдоль всего тела - 4 гиподермальных валика: спинной, два боковых и брюшной. Под гиподермой располагаются тяжи продольных мышц.
Внутри кожно-мускульного мешка расположена первичная полость тела (псевдоцель), где помещаются все внутренние органы. Протоцель заполнена жидкостью под давлением и выполняет роль гидроскелета - опорную функцию. Первичная полость тела выполняет также функцию транспортную (распределительную) - переносит вещества от кишечника к рабочим органам, в том числе - мышцам.
Начинается ротовой полостью, далее продолжается в пищевод, затем в переднюю, среднюю и заднюю кишку с анальным отверстием, открывающимся с брюшной стороны на заднем конце тела. Сквозная пищеварительная система обеспечивает возможность непрерывного питания.
Дыхание осуществляется всей поверхностью тела, у паразитов - анаэробное (бескислородное) дыхание.
Жидкие продукты обмена веществ удаляются через органы выделения - видоизмененные протонефридии (без мерцательного "пламени", которое было у плоских червей). По бокам тела имеются экскреторные боковые каналы, которые в передней части тела соединяются в общий выводной проток. Этот проток заканчивается порой на головном конце тела с брюшной стороны.
Помимо этого в коже имеется много мелких эпидермальных желез, связанных с экскреторными боковыми каналами. Функцией выделения обладают также особые клетки - фагоцитарные. Они накапливают в себе продукты обмена веществ и инородные тела, попавшие в полость тела. Фактически они не выделяют, а накапливают эти вещества, таким образом, удаляют их из организма.
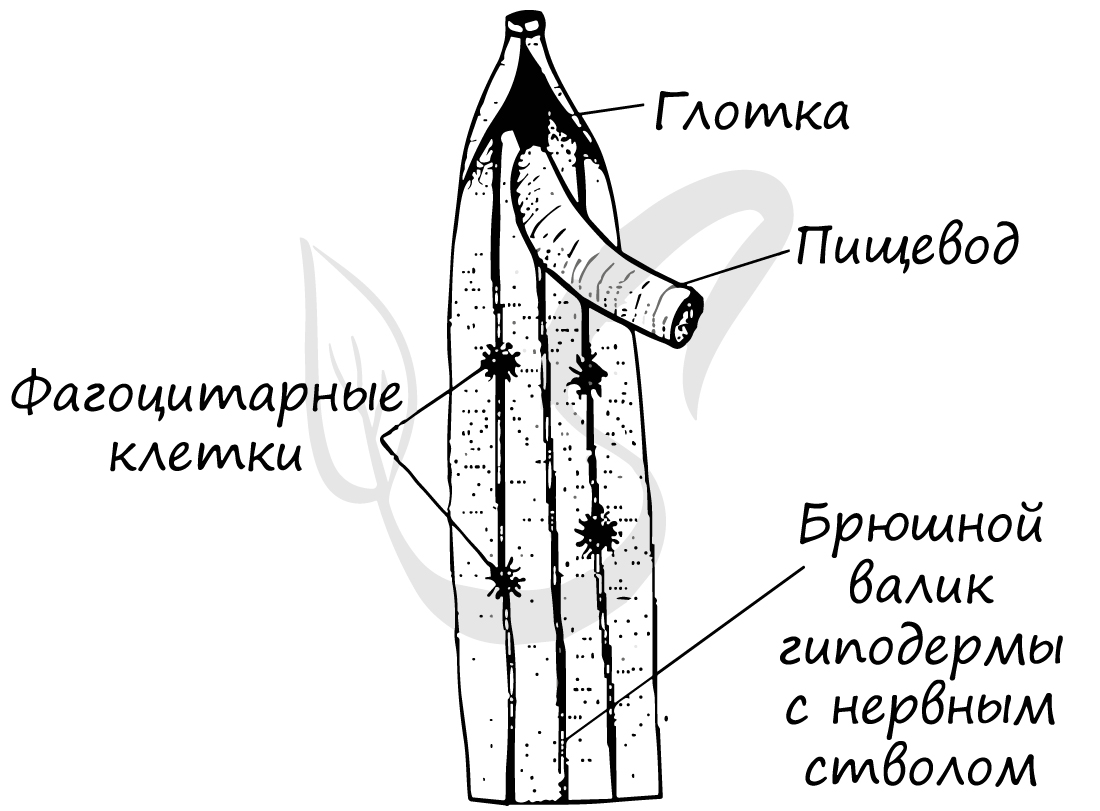
Нервная система круглых червей стволового (лестничного) типа, также называемая - ортогон. Состоит из окологлоточного нервного кольца, от которого отходят нервные стволы, из которых наиболее выделяются брюшной и спинной. Стволы соединены друг с другом поперечными тяжами - комиссурами.
Строение окологлоточного нервного кольца отражает единство всей его структуры - по сути это один большой круговой ганглий (нервный узел).
Круглые черви раздельнополы. Развитие, как правило, непрямое, без метаморфоза - молодая особь похожа на взрослую, только меньше ее в размерах. У части паразитических форм присутствует стадия инвазионной личинки.
Женские половые органы представлены влагалищем, яичником, яйцеводом и маткой. Возможен парный набор женских половых органов, но влагалище всегда одно.
Мужская половая система состоит из семенников с семяпроводами, и единственного семяизвергательного канала. Спермии круглых червей (спермий отличается от сперматозоида отсутствием жгутика) имеют разнообразное строение и отличаются амебоидной подвижностью.
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Информация
Добавить в ЗАКЛАДКИ| Поделиться: |
Анаэробное дыханиеДЫХАНИЕ КЛЕТОЧНОЕ — химический процесс распада органических молекул в клетке с выделением энергии, необходимой для жизнедеятельности. У большинства организмов разложение глюкозы в присутствии кислорода до углекислого газа и воды. Некоторые микробы способны получать достаточное количество энергии в ходе частичного разложения органики и отсутствия кислорода с образованием иных конечных продуктов (брожение, анаэробное дыхание).[ . ] Дыхание анаэробов идет без участия кислорода. Они получают необходимую энергию расщеплением сложной молекулы органического вещества на более простые. При этом выделяется гораздо меньше энергии , чем при кислородном дыхании. Примером анаэробного дыхания может служить процесс брожения глюкозы.[ . ] Анаэробное дыхание служит основой жизнедеятельности главным образом бактерий, дрожжей, плесневых грибов и др., хотя как звено метаболизма оно может встречаться и в некоторых тканях высших животных. Наиболее характерные примеры анаэробного дыхания:, .образование метана метановыми бактериями за о-чет разложения органического соединения или восстановления угля или карбонатов, образование сероводорода сульфатвосстанавливающими бактериями ( в частности,в Черном море), винное брожение. Анаэробное дыхание выделяет меньше энергии,чем аэробное. Полегают, что первичный мир живого имел анаэробные формы, на оонове которых позднее образовался аэробный мир.[ . ] Анаэробное дыхание — реакции распада глюкозы без участия кислорода.[ . ] Итак, дыхание — процесс гетеротрофный, приблизительно уравновешивающий автотрофное накопление органического вещества. Различают аэробное, анаэробное дыхание и брожение.[ . ] Этап неполного окисления (анаэробное дыхание или брожение). Неполному окислению могут подвергаться глюкоза, жирные кислоты, аминокислоты. При этом главным источником энергии в клетке является глюкоза. При бескислородном окислении одной молекулы глюкозы (процесс гликолиза) из двух молекул АДФ образуются две молекулы АТФ. В процессе гликолиза для нужд клетки извлекается не более 10% энергии.[ . ] С точки зрения выхода энергии, анаэробное дыхание - крайне неэффективный процесс, так как значительная часть энергии, содержащейся в шестиугольном сахаре, все еще остается в связях молекул спирта.[ . ] Схема 1. Транспорт электронов при дыхании в различных типах анаэробного получения энергии (брожение и анаэробное дыхание).[ . ] Микроорганизмы, имеющие факультативно-анаэробное дыхание, в своих клетках содержат, кроме дегидраз, еще оксидазы и ферменты, активирующие кислород, т. е. ферменты, свойственные и аэробным микробам. Дрожжи относятся к группе факультативно-анаэробных микроорганизмов, т. е. им свойственно и анаэробное и аэробное дыхание, но последнее выражено слабее. При анаэробном дыхании дрожжи расходуют на дыхание значительно больше энергетического материала (сахара), чем при аэробном дыхании.[ . ] Критическая концентрация кислорода для начала анаэробного дыхания равна 3. 10% во внешней атмосфере.[ . ] Бактерии ВезиШиЬЬпо — экологически важный пример анаэробного дыхания (тип 2); эти бактерии восстанавливают 804 в глубоких отложениях и в бескислородных водах, например б Черном море, до газообразного НгЭ. Этот газ может подеяться в верхние слои отложений или в поверхностные воды, где его используют другие организмы (например, фотосинтезирующие бактерии). К общеизвестным организмам, использующим брожение (тип 4), относятся, разумеется, дрожжи. Они имеют большую практическую ценность для человека, но, кроме того, в изобилии встречаются в почве, где играют ключевую роль в разложении растительных остатков.[ . ] Величина ДК зависит и от других причин. В некоторых тканях из-за затрудненного доступа кислорода наряду с аэробным происходит анаэробное дыхание, не сопровождающееся поглощением кислорода, что приводит к повышению значения ДК. Величина коэффициента обусловлена также полнотой окисления дыхательного субстрата. Если, кроме конечных продуктов, в тканях накапливаются менее окисленные соединения (органические кислоты), то ДК< 1.[ . ] Таким образом, на основании вышеизложенного можно сделать вывод, что необходимую энергию микроорганизмы получают или в процессе дыхания, заключающегося в использовании кислорода и в полном превращении органических соединений в углекислоту и воду, или в результате брожения, которое происходит без участия кислорода и заканчивается получением продуктов неполного окисления. В. И. Палладии считал, что между дыханием и брожением, которое он считал анаэробным дыханием, нет принципиальной разницы.[ . ] Метановое брожение известно давно (мерцающие огоньки на кладбищах, выделение болотного газа). Оно происходит при разложении продуктов, содержащих углеводы. Это способ анаэробного дыхания определенных групп бактерий, которые из углеводов органической массы образуют метан (СН4) - 65%, углекислый газ (СОг) - 30% и незначительное количество других газов: сероводород (Н35) - 1%, азот, кислород, водород и закись углерода. При этом выделяется энергия, которая превращается в тепловую и нагревает субстрат.[ . ] Рост грибов и выделение ими углекислоты зависят от давления кислорода в атмосфере и от температуры. При меньшем давлении кислорода чем 1,5 атм и температуре 17,5° С гриб перестает расти и его обмен приобретает анаэробный характер. Нижняя граница аэробного дыхания зависит от температуры: при 29,5° С обмен веществ уже меняется при давлении кислорода 1,5 атм. В условиях анаэробного дыхания выделение углекислоты прямо пропорционально давлению кислорода. В таких условиях изменяется весь обмен веществ гриба, весь набор его ферментов.[ . ] Углеводы являются основным продуктом фотосинтеза, на их основе в процессе обмена веществ в растительном организме формируются белки, жиры, нуклеиновые кислоты и другие соединения. Углеводы - основной источник для аэробного и анаэробного дыхания клеток; источник энергии для возобновления вегетации. Обычно растение содержит большой набор разнообразных углеводов. В процессе вегетации соотношение растворимых и нерастворимых форм изменяется. В молодых растениях преобладают моно- и дисахариды, в период созревания увеличивается содержание крахмала, целлюлозы, т.е. нерастворимых форм.[ . ] В зависимости от вида сырья плоды поступают на хранение в деревянных ящиках, ящичных поддонах, контейнерах, насыпью. При этом желательно не укладывать сырье (особенно насыпью) очень высоким слоем, иначе будет затруднен доступ воздуха к отдельным плодам. В этом случае процесс нормального (аэробного) дыхания нарушится и наступит так называемое интрамолекулярное или анаэробное дыхание, протекающее по приведенной выше схеме спиртового брожения и приводящее сырье к порче. Поэтому сырье, особенно нежной консистенции, укладывают в ящики-клетки штабелями высотой до 2 м, оставляя между отдельными штабелями проходы. При таком хранении имеется достаточный доступ воздуха и дыхание протекает нормально.[ . ] Однако русский ученый В. О. Таусон показал, что для многих синтетических реакций, происходящих в живой протоплазме микробов, дыхательный процесс как источник энергии не нужен. Выделение углекислоты в ряде процессов также не связано с поглощением кислорода и с полным окислением питательных веществ, т. е. с процессом дыхания, а является результатом процесса, получившего название анаэробного дыхания, или брожения.[ . ] Получение энергии путем разложения части биомассы характерно для всех живых организмов, независимо от того, как эта биомасса появляется. Под разложением в живых организмах понимают любые биологические окисления, дающие энергию. При этом окислителем (акцептором электронов) может служить газообразный кислород (аэробное дыхание), какое-либо неоргавичеокое или органическое соединение (анаэробное дыхание),а также самоокисляемое соединение (брожение - разновидность анаэробного дыхания).[ . ] Применение вермикулита в качестве упаковочного материала значительно улучшает хранение картофеля, моркови и других овощей. Это объясняется прежде всего его влагопоглощающими свойствами, благодаря которым на поверхности объектов хранения никогда не бывает капельно-жидкой влаги. На сухой тургесцентной поверхности не происходит развития грибных гнилей и бактериозов. Размножению грибов препятствует также щелочная реакция консерванта. Вермикулит, обладая адсорбционными свойствами, способствует удалению из окружающей овощи атмосферы ацетальдегида, этилового спирта и других продуктов анаэробного дыхания. Этим предупреждаются функциональные расстройства (физиологические болезни) картофеля и овощей. Сыпучесть и легкость проникновения вермикулита между отдельными экземплярами овощей ограничивает контакт между больными и здоровыми клубнями или корнеплодами, следовательно, исключается гнездовое загнивание продукции. Этот сыпучий минерал может играть роль носителя фунгицидов и бактерицидов, а также поддерживать нужную концентрацию влаги в атмосфере.[ . ] Читайте также:
 Пожалуйста, не занимайтесь самолечением!При симпотмах заболевания - обратитесь к врачу. Пожалуйста, не занимайтесь самолечением!При симпотмах заболевания - обратитесь к врачу.
|