
Сопряженная эволюция паразита и хозяина
Паразитизм – форма взаимоотношения между двумя организмами разных видов, про котором один использует другого в качестве источника питания и среды обитания, причем оба организма находятся в антагонистических отношениях, но ,в отличие от хищничества, паразит не убивает своего хозяина, а паразитирует на нем более или менее продолжительный период времени.
Сопряженная эволюция паразита и хозяина характеризуется совершенствованием механизмов противопаразитарной борьбы у хозяина и повышением приспособляемости паразитов.
У хозяев вырабатывается целый ряд защитных реакций , которые делятся на специфические и неспецифические.
· к неспецифическим относится иммунный ответ организма, т. е. возникновение биохимических реакций, которые сдерживают массовое развитие паразитов;
· сбрасывание зараженных частей (это особенно характерно для растений-хозяев, которые сбрасывают сильно зараженные листья). В этом случае паразиты продолжают жить уже как детритофаги;
· выработка устойчивости к влиянию паразитов за счет быстрого роста здоровых тканей взамен пораженных (это имеет место при поедании тканей растений тлями);
· уменьшение плотности популяций хозяев, что снижает вероятность распространения паразита и заражения им. Зараженные животные менее подвижны и становятся более легкой добычей хищников, таким образом, снижая долю зараженных особей в популяции;
· формирование гетерогенных популяций хозяев, в составе которых есть экотипы, устойчивые к паразитам. Эти экотипы являются основой адаптивной селекции на повышение устойчивости культурных растений к грибковым заболеваниям.
У паразитов также в ходе совместной эволюции вырабатываются защитные механизмы
· инцистирование и инкапсуляция, капсулы вокруг паразита формируется из тканей хозяина. Такая защитная изоляция возможна лишь у покоящихся стадий паразитов, которые характеризуются минимальным уровнем обмена веществ.
· Локализация в тканях наиболее изолированных от действия иммунной системы. Чтобы избежать атаки защитных сих хозяина, некоторые паразиты поселяются в наиболее защищенных от воспалительных реакций тканях. Например, метацеркарии некоторых трематод.
· Иммуносупрессия – способность подавлять действие иммунной защиты хозяина.
· Изменение состава антигенов.
· Молекулярная мимикрия – паразит покрывает себя веществами, образованными хозяином, благодаря чему скрывается от распознавания средствами его иммунной системы. К такой мимикрии способны церкарии шистосом.
В процессе адаптации к своим хозяевам у паразитов происходит морфофизиологический регресс органов, функции которых не востребованы. Наряду с этим у паразитов развиваются физиологические механизмы и структурные приспособления, необходимые для существования в организме хозяина.
· Изменяется форма тела. У эктопаразитов тело сплющено в дорсо - вентральном направлении, что облегчает удержание на теле хозяина. У блох же тело сплющен ос боков. Внутриполостные паразиты имеют компактное тело.
· Размеры обычно коррелируют с размерами соответствующих групп свободноживущих хозяев. В некоторых случаях паразитизм приводит к увеличению размеров тела. Но размеры паразита всегда много меньше размеров хозяина.
· Большим изменениям подвергается пищеварительная система, у некоторых она совсем отсутствует (ленточные черви). У кровососущих она имеет большую емкость и способна вместить значительное количество крови.
· Разнообразные приспособления для удержания в или на организме хозяина. Различают разнообразные крючки и присоски. Крючки встречаются, начиная с простейших (например, у грегарин) вплоть до членистоногих. Помимо органов присосок и крючков органами прикрепления служат различные стилеты, стрекательные и липкие нити, стебельки и другие приспособления.
Вместе с морфологическими изменениями у паразитов происходят и физиологические адаптации к паразитическому образу жизни
· Внутриполостные и тканевые паразиты приобрели способность к анаэробному дыханию
· Кишечные паразиты приобрели способность вырабатывать антиферменты, которые защищают их от пищеварительных ферментов хозяина.
· Некоторые паразиты (личинки шистосом и анкилостомид) стали вырабатывать ферменты, с помощью которых получили возможность проникать через кожные покровы в организм хозяина.
· Большая плодовитость, гермафродитизм также являются приспособлением к паразитическому образу жизни.
Для естественных экосистем формирование экологического равновесия между популяциями паразитов и их хозяев — нормальное явление. Во многих случаях проявляется закономерность: плотность популяций обоих видов изменяется циклически, но пики плотности паразитов запаздывают по отношению к пикам плотности хозяев. Ситуация изменяется в антропогенных экосистемах (особенно в сельскохозяйственных), где заражение паразитами может привести к существенному падежу скота.
Основная особенность отрицательного взаимодействия популяций заключается в том, что при их синхронной эволюции в стабильной экосистеме степень отрицательного влияния уменьшается. Иными словами, естественный отбор стремится уменьшить отрицательные влияния или вообще устранить взаимодействие популяций, поскольку продолжительное и сильное подавление популяции добычи или хозяина популяцией хищника илипаразита может привести к уничтожению одной из них или обеих.
- Материально - энергетический баланс производства. Потоки энергии в производственных процессах. Энергетическая эффективность деятельности. Функционально-стоимостной анализ как метод оптимизации.
Для описания технологических процессов широко используют материальные и энергетические балансы.
Материальный баланс — это отражение закона сохранения массы веществ в условиях производства. Согласно этому закону, масса исходных веществ (сырья), поступивших для участия в технологическом процессе, равна массе веществ (продуктов и отходов), образовавшихся в результате осуществления технологического процесса. Как правило, материальный баланс составляется на единицу целевого продукта.
Энергетический баланс — это количественное выражение закона сохранения энергии в ходе осуществления технологического процесса. Согласно этому закону, количества энергии, потребленной и выделяющейся в ходе процесса, равны. Как и материальный, энергетический баланс составляется на единицу целевого продукта.
Материальные и энергетические балансы имеют большое значение для анализа и эффективного осуществления производственного процесса. С их помощью устанавливают фактический выход продукции, коэффициенты полезного использования энергии, расходы и потери сырья, топлива и других материалов. По цифровым данным технологических балансов принимаются организационные и технические решения по совершенствованию работы оборудования, максимальному использованию, утилизации или регенерации материальных и энергетических ресурсов. Балансы весьма важны для составления рациональных схем технологического процесса и установления оптимальных размеров, мощности и производительности оборудования. Данные технологических балансов и выводы из их анализа широко используются технологами и экономистами при проектировании новых и улучшении работы существующих предприятий.
ФУНКЦИОНАЛЬНО-СТОИМОСТНЫЙ АНАЛИЗ (ФСА) — это практический метод комплексного системного исследования объектов, направленный на обеспечение минимальных затрат на производство, реализацию или эксплуатацию при обеспечении необходимых потребительских свойств.
ФСА от других методов повышения эффективности производства отличает совокупность принципов, которая включает плановый характер проведения ФСА, комплексный подход, системный подход, функциональный подход, принципы соответствия значимости функций и затрат на их осуществление, соответствия фактического значения параметра требуемому уровню, активизации творческого мышления, коллективного труда, междисциплинарного подхода и др.
Дата публикования: 2015-02-03 ; Прочитано: 1100 | Нарушение авторского права страницы
studopedia.org - Студопедия.Орг - 2014-2020 год. Студопедия не является автором материалов, которые размещены. Но предоставляет возможность бесплатного использования (0.003 с) . 
Паразитизм, как явление, возник на том этапе развития орга-нического мира, когда на Земле появились достаточно разнообразные живые организмы, отличающиеся размерами тела и образом питания. С появлением разнообразных форм жизни возникли первые биоценозы. Укрепление и расширение экологических связей в биоценозах привели к возникновению симбиозов. Симбиозом является любая форма совместного существования особей разных видов. Паразитизм эволюционно возник в процессе расширения и укрепления пищевых и пространственных связей симбионтов (см. табл.).
Симбиоз - любое совместное существование особей разных видов.
Воздействие паразита на организм хозяина:
- механическое повреждение органов и тканей;
- поглощение и нарушение всасывания питательных, минеральных веществ и витаминов;
- нарушение обмена веществ;
- токсическое действие;
- иммунодепрессивное действие;
- изменение поведения хозяина.
Хозяин отвечает на внедрение паразита различными реакциями, которые проявляются на клеточном, тканевом и организмен- ном уровнях. Между паразитом и хозяином устанавливаются особые взаимоотношения (система: паразит-хозяин), при которых паразит, благодаря различным адаптациям, длительное время использует хозяина, не убивая его.
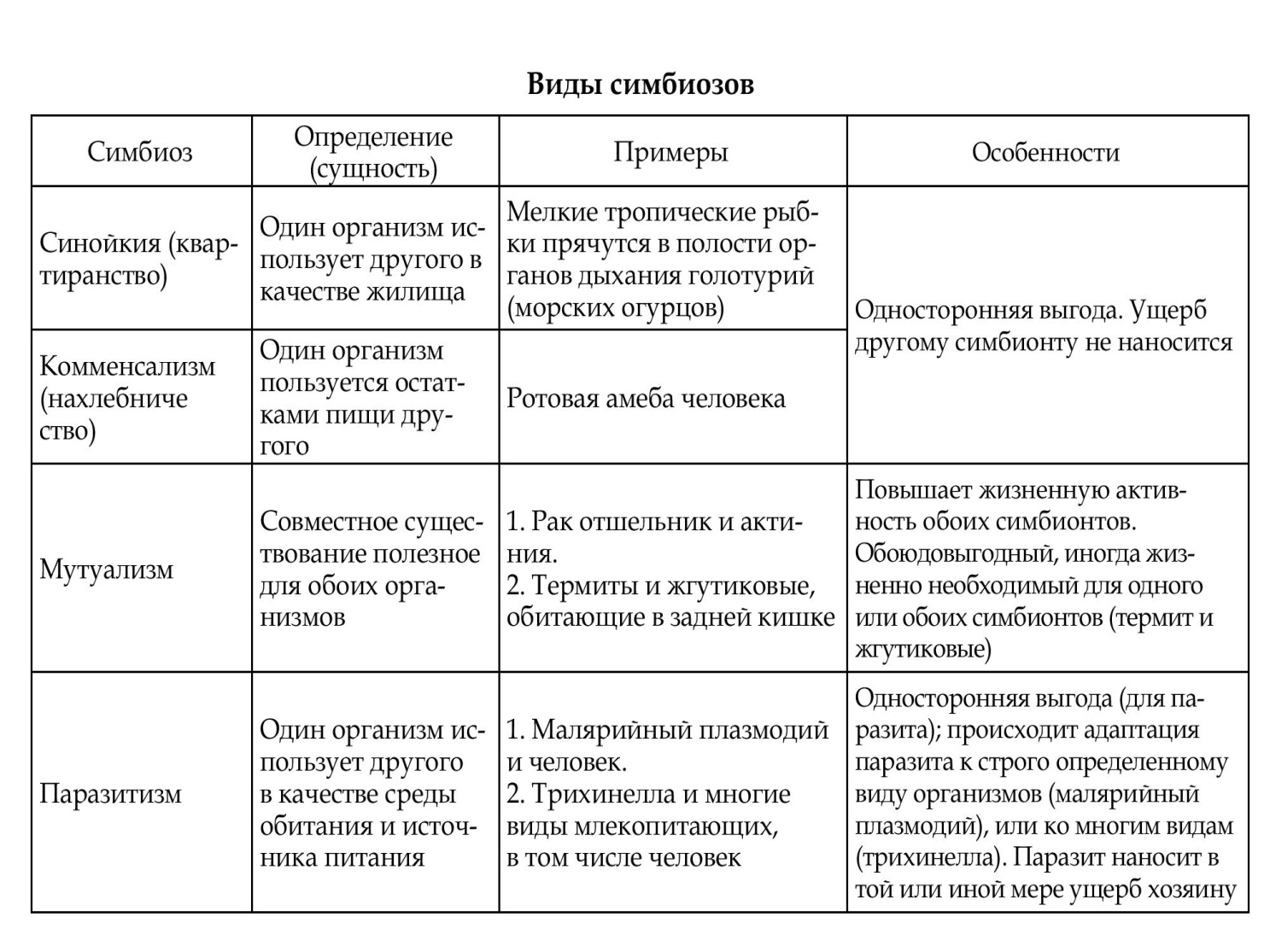
Классификация паразитов
Формы паразитизма по особенностям:
облигатный - организм, в жизненном цикле которого стадия паразитирования обязательна (сосальщики, вши и др.);
факультативный - организм, который может жить и питаться свободно во внешней среде, но вступая в контакт с восприимчивым организмом, может переходить к паразитизму (амебы группы Umax, конская пиявка); ложный - свободноживущий организм, при случайном проникновении в организм другого вида может некоторое время сохранять жизнеспособность (личинки сырной мухи в кишечнике человека).
Продолжительность контакта с хозяино:
постоянный (стационарный) - паразит, весь жизненный цикл которого связан с организмом хозяев (вши); временный - организм, который вступает в паразитарные отношения с хозяином только на время питания (иксодовые клещи, блохи, комары).
Видовая специализация:
паразитов: эвриксенный (греч. eurys - широкий; xenon - хозяин) - па-разит, который имеет широкий круг хозяев (трихинелла, комары, иксодовые клещи);
стеноксенный (греч. stenos - узтш) - паразит преимущест-венно одного вида хозяев, но способный паразитировать и на некоторых других видах (крысиная блоха); моноксенный (греч. monos - один) - паразит, который имеет хозяев только одного вида (аскарида человеческая, криво- головка, острица, вошь человеческая головная и др.).
Медицинское значение:
переносчики - кровососущие членистоногие, передающие возбудителей заболеваний (инфекционных и инвазионных) от одного организма к другому (комары, иксодовые клещи, москиты, блохи и др.); возбудители - паразиты, которые вызывают инвазионные заболевания (лейшмании, трипаносомы, гельминты), инфестации (чесоточный клещ, вши, личинки вольфартовой мухи).
Локализация:
эктопаразит - организм, который паразитирует на по-кровах тела хозяина (комары, вши, иксодовые клещи и др.);
эндопаразит - организм, который паразитирует внутри организма хозяина (схема).
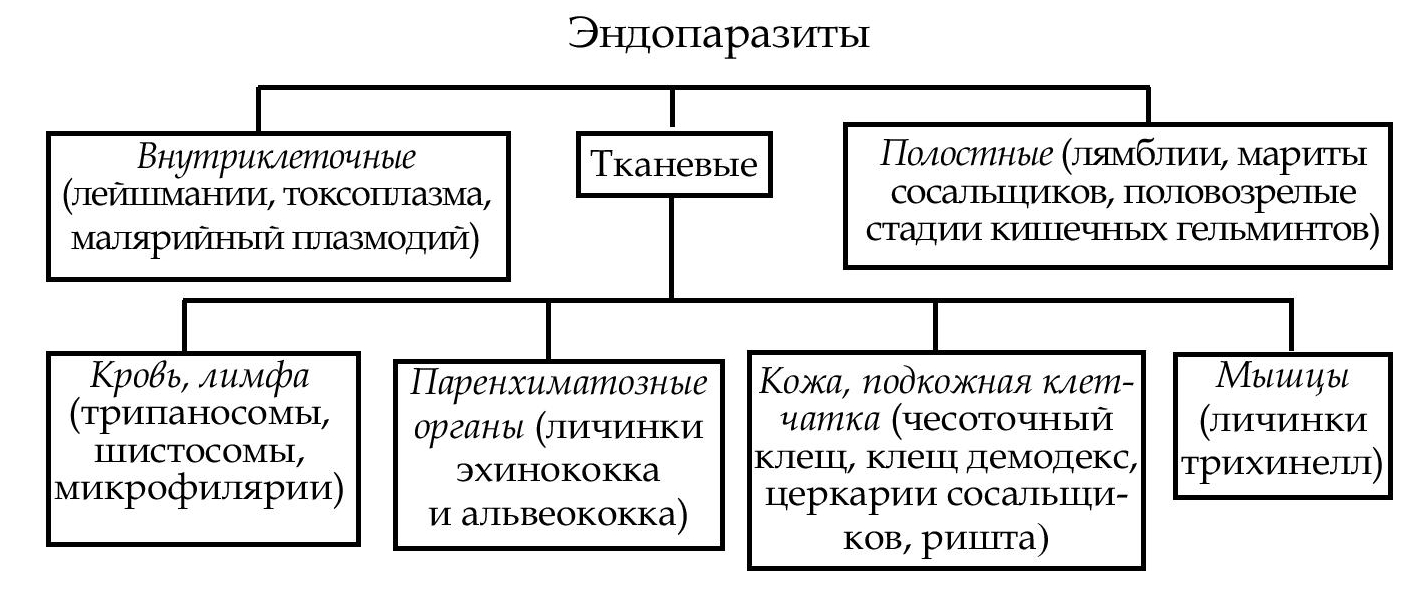
В зависимости от того, кто болеет и является источником возбу-дителя болезни, паразитарные заболевания делятся следующим образом:
- антропонозы- заболевания, в цикле развития возбудителей которых человек является обязательным звеном (малярия, лямблиоз, амебиаз, энтеробиоз, филяриоз и др.);
- зоонозы - болезни, поражающие животных и человека. Ис-точником заражения в основном являются животные, а человек - как правило, тупиковое звено в развитии паразита (трихинеллез, токсоплазмоз, эхинококкоз и др.).
Классификация паразитарных болезней в соответствии с систематическим положением паразитов
Протозоозы- паразитарные болезни, возбудители которых от-носятся к типу Простейшие (амебиаз, лейшманиоз, токсоплазмоз).
Гельминтозы- паразитарные болезни, возбудители которых относятся к типам Плоские и Круглые черви.
Гельминтозычеловека
Трематодозы- возбудители относятся к классу Сосальщики.
Цестодозы - возбудители относятся к классу Ленточные Черви.
Нематодозы - возбудители относятся к классу Собственно Круглые Черви.
По биологическим особенностям циклов развития среди гельминтов выделяются группы геогельминтов и биогельминтов. По особенностям передачи инвазии выделяют контактно-передавае- мых гельминтов.
Геогельминты - такие гельминты, развитие личиночных стадий которых происходит во внешней среде. Промежуточный хозяин отсутствует (аскарида, власоглав, кривоголовки).
Биогельминты - такие гельминты, в жизненном цикле которых происходит смена хозяев (сосальщики, вооруженный и невооруженный цепни, лентец, ришта) или развитие всех стадий происходит в одном организме без выхода во внешнюю среду (трихинелла, карликовый цепень).
Контактно-передаваемые (контагиозные) гельминты - особая группа гельминтов, инвазионная стадия которых может попадать в организм человека непосредственно при контакте с больным (карликовый цепень, острица). При контактно-передаваемых гельминтозах возможна аутореинвазия.
Инфестации - паразитарные болезни, возбудители которых относятся к типу Членистоногие (чесотка, педикулез, демодекоз и др.).

Инвазионная стадия - стадия развития паразита, внедрение которой в организм хозяина способно вызвать заболевание.

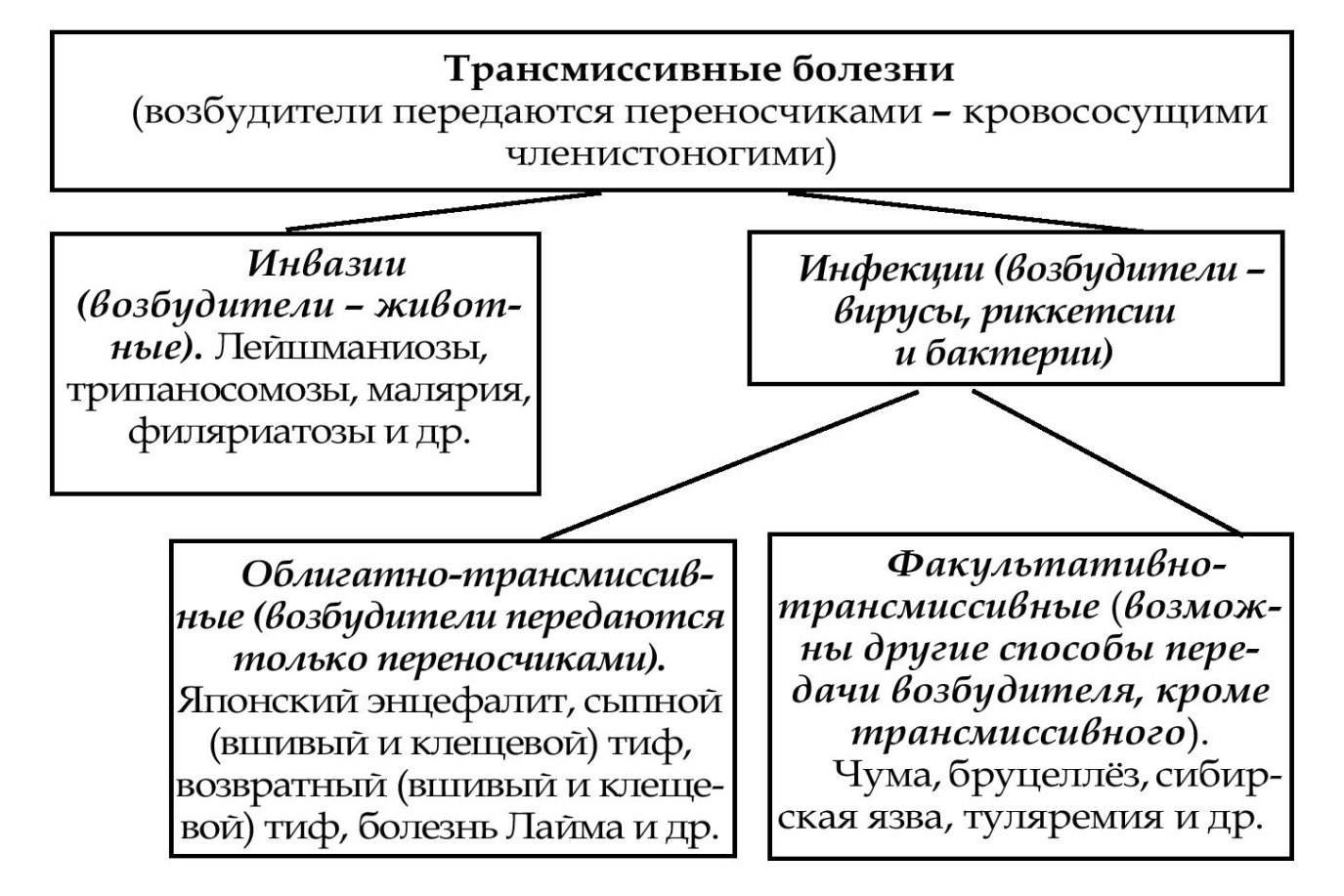
Варианты трансмиссивной передачи возбудителей переносчиками
Инокуляция(от лат. inoculatio - прививка) - введение возбудителя болезни переносчиком в ранку с помощью ротового аппарата.
• Специфическая - переносчик специфический (передача возбудителей малярии комарами рода Anopheles, чумы - блохами, клещевого энцефалита - иксодовыми клещами).
• Неспецифическая (механическая) - переносчик неспецифический (передача возбудителей сибирской язвы, туляремии, бруцеллеза осенней жигалкой, слепнями и др.).
Контаминация (от лат. contaminatio - загрязнение) - возбудитель наносится переносчиком на покровы тела, а затем через микротравмы проникает в организм хозяина (передача вшами возбудителей сыпного и возвратного тифов).
Классификация переносчиков
Переносчик - кровососущие членистоногие, передающие возбудителей болезней от одного организма к другому.
• Специфический - переносчик, в организме которого возбудитель проходит определенные стадии развития или размножается (комары рода Anopheles для малярийного плазмодия, москиты для лейшманий, мухи цеце и поцелуйные клопы для трипаносом определенных видов, иксодовые клещи для вируса клещевого энцефалита, блохи для бактерий чумы).
• Неспецифический - переносчик, выполняющий функцию механического переноса возбудителя болезни без развития и размножения последнего (слепни, осенние жигалки и иксодовые клещи для возбудителей туляремии, бруцеллеза, сибирской язвы).
Пример: в жизненном цикле лентеца широкого:
- окончательный хозяин (плотоядные животные и человек);
- промежуточный хозяин (низшие ракообразные);
- дополнительный хозяин (пресноводные рыбы, питающиеся зоопланктоном);
- резервуарный хозяин (хищные рыбы).
Хозяин-прокормитель (для эктопаразитов) - организм, который служит только для питания паразита (может стать резервуаром возбудителя болезни в очаге и источником возбудителя). Например, лось является хозяином-прокормителем для иксодовых клещей.

Учение академика Е. Н. Павловского о природной очаговости болезней человека
Суть учения состоит в том, что в природных биогеоценозах независимо от человека происходит циркуляция возбудителей болезней. Такие болезни получили название природно-очаго- вых. Природно-очаговые болезни существуют в определенных биогеоценозах, их возбудители циркулируют среди диких животных. Люди, попадающие в эти биогеоценозы, могут подвергаться заражению. Циркуляция возбудителей природно-очаго- вых болезней может происходить как с участием переносчиков (природно-очаговые трансмиссивные болезни), так и без участия переносчиков (природно-очаговые нетрансмиссивные болезни).
К природно-очаговым трансмиссивным болезням относятся: трипаносомозы, некоторые формы лейшманиоза, таежный энцефалит, японский энцефалит, болезнь Лайма, чума, туляремия, клещевые возвратный и сыпной тифы.
К природно-очаговым нетрансмиссивным болезням относятся: токсоплазмоз, трихинеллез, дифиллоботриоз, эхинококкоз, альвеококкоз, описторхоз, парагонимоз, шистосомозы. В природных очагах нетрансмиссивных заболеваний циркуляция возбудителей между хозяевами осуществляется без участия переносчиков через факторы среды.
Природный очаг - это участок биогеоценоза, на котором циркуляция возбудителя происходит среди диких животных независимо от человека.
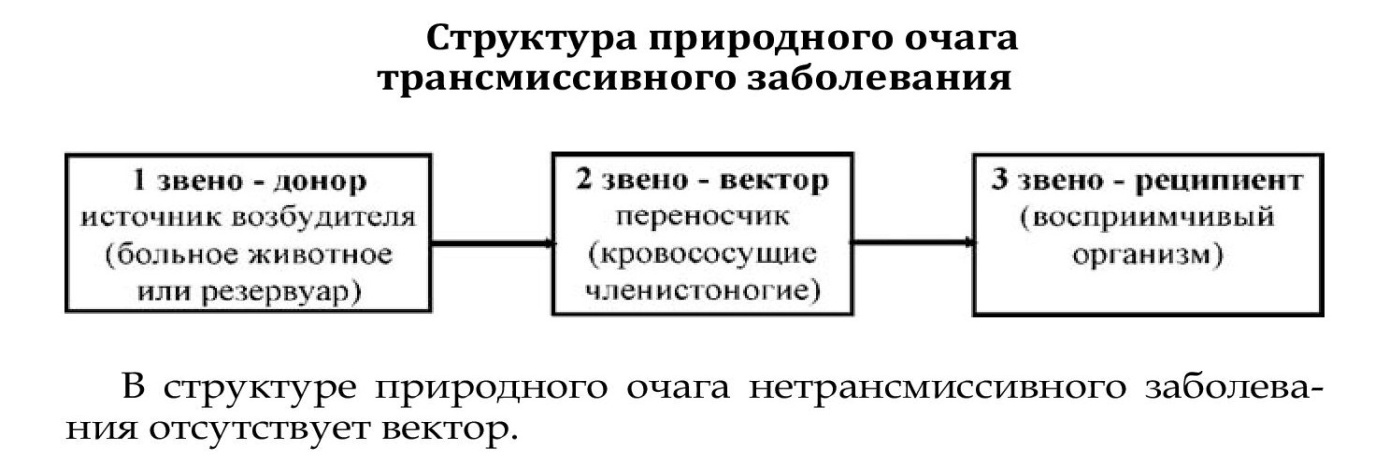
В структуре природного очага нетрансмиссивного заболевания отсутствует вектор.
Классификация природных очагов По количеству хозяев:
- моногостальный - резервуаром является один вид животных. Формируется в тех случаях, если возбудитель адаптирован только к одному виду хозяев или на определенной территории не обитают другие виды животных, которые могут быть хозяевами паразита (природный очаг африканского трипаносомоза, резервуар - антилопы);
- полигостальный - резервуаром являются несколько видов животных (природный очаг клещевого энцефалита, резервуары - птицы, млекопитающие).
По количеству переносчиков:
- моновекторный - возбудители передаются только одним видом переносчиков. Определяется видовым составом переносчиков в конкретном биоценозе (в определенном очаге таежного энцефалита обитает только один вид иксодовых клещей);
- поливекторный - возбудители передаются различными видами переносчиков (природный очаг туляремии - переносчики: различные виды комаров, слепней, иксодовых клещей).
По характеру освоения ландшафта (уровню антропопрессии):
- антропургический - циркуляция возбудителя происходит среди домашних и диких животных. Возникает при освоении человеком территории природного очага (такой характер могут приобретать очаги японского энцефалита, кожного лейшманио- за, клещевого возвратного тифа, трипаносомоза и др.);
- синантропный - циркуляция возбудителя происходит среди синантропных и домашних животных (очаги трихинеллеза, ток- соплазмоза и др.).
Природно-очаговые болезни характерны преимущественно для диких животных. Однако антропопрессия и урбанизация создают условия для распространения возбудителей этих заболеваний среди синантропных животных и людей. Так возникают ант- ропургические, а затем и синантропные очаги болезней, которые могут представлять значительную эпидемическую опасность. Санкт-Петербургский мегаполис имеет в своей черте многочисленные лесопокрытые площади природно-антропогенного происхождения (скверы, сады, лесопарки и др.). Значительная часть этих территорий заселена иксодовыми клещами, среди которых доминирует таежный клещ. Ежегодно сотни горожан подвергаются нападению клещей и рискуют заразиться опасными транс-миссивными заболеваниями, природные очаги которых находятся на территории Санкт-Петербурга и Ленинградской области.
Паразитизм – форма взаимоотношения между двумя организмами разных видов, про котором один использует другого в качестве источника питания и среды обитания, причем оба организма находятся в антагонистических отношениях, но ,в отличие от хищничества, паразит не убивает своего хозяина, а паразитирует на нем более или менее продолжительный период времени.
Сопряженная эволюция паразита и хозяина характеризуется совершенствованием механизмов противопаразитарной борьбы у хозяина и повышением приспособляемости паразитов.
У хозяев вырабатывается целый ряд защитных реакций , которые делятся на специфические и неспецифические.
к неспецифическим относится иммунный ответ организма, т. е. возникновение биохимических реакций, которые сдерживают массовое развитие паразитов;
сбрасывание зараженных частей (это особенно характерно для растений-хозяев, которые сбрасывают сильно зараженные листья). В этом случае паразиты продолжают жить уже как детритофаги;
выработка устойчивости к влиянию паразитов за счет быстрого роста здоровых тканей взамен пораженных (это имеет место при поедании тканей растений тлями);
уменьшение плотности популяций хозяев, что снижает вероятность распространения паразита и заражения им. Зараженные животные менее подвижны и становятся более легкой добычей хищников, таким образом, снижая долю зараженных особей в популяции;
формирование гетерогенных популяций хозяев, в составе которых есть экотипы, устойчивые к паразитам. Эти экотипы являются основой адаптивной селекции на повышение устойчивости культурных растений к грибковым заболеваниям.
У паразитов также в ходе совместной эволюции вырабатываются защитные механизмы
инцистирование и инкапсуляция, капсулы вокруг паразита формируется из тканей хозяина. Такая защитная изоляция возможна лишь у покоящихся стадий паразитов, которые характеризуются минимальным уровнем обмена веществ.
Локализация в тканях наиболее изолированных от действия иммунной системы. Чтобы избежать атаки защитных сих хозяина, некоторые паразиты поселяются в наиболее защищенных от воспалительных реакций тканях. Например, метацеркарии некоторых трематод.
Иммуносупрессия – способность подавлять действие иммунной защиты хозяина.
Изменение состава антигенов.
Молекулярная мимикрия – паразит покрывает себя веществами, образованными хозяином, благодаря чему скрывается от распознавания средствами его иммунной системы. К такой мимикрии способны церкарии шистосом.
В процессе адаптации к своим хозяевам у паразитов происходит морфофизиологический регресс органов, функции которых не востребованы. Наряду с этим у паразитов развиваются физиологические механизмы и структурные приспособления, необходимые для существования в организме хозяина.
Изменяется форма тела. У эктопаразитов тело сплющено в дорсо - вентральном направлении, что облегчает удержание на теле хозяина. У блох же тело сплющен ос боков. Внутриполостные паразиты имеют компактное тело.
Размеры обычно коррелируют с размерами соответствующих групп свободноживущих хозяев. В некоторых случаях паразитизм приводит к увеличению размеров тела. Но размеры паразита всегда много меньше размеров хозяина.
Большим изменениям подвергается пищеварительная система, у некоторых она совсем отсутствует (ленточные черви). У кровососущих она имеет большую емкость и способна вместить значительное количество крови.
Разнообразные приспособления для удержания в или на организме хозяина. Различают разнообразные крючки и присоски. Крючки встречаются, начиная с простейших (например, у грегарин) вплоть до членистоногих. Помимо органов присосок и крючков органами прикрепления служат различные стилеты, стрекательные и липкие нити, стебельки и другие приспособления.
Вместе с морфологическими изменениями у паразитов происходят и физиологические адаптации к паразитическому образу жизни
Внутриполостные и тканевые паразиты приобрели способность к анаэробному дыханию
Кишечные паразиты приобрели способность вырабатывать антиферменты, которые защищают их от пищеварительных ферментов хозяина.
Некоторые паразиты (личинки шистосом и анкилостомид) стали вырабатывать ферменты, с помощью которых получили возможность проникать через кожные покровы в организм хозяина.
Большая плодовитость, гермафродитизм также являются приспособлением к паразитическому образу жизни.
Для естественных экосистем формирование экологического равновесия между популяциями паразитов и их хозяев — нормальное явление. Во многих случаях проявляется закономерность: плотность популяций обоих видов изменяется циклически, но пики плотности паразитов запаздывают по отношению к пикам плотности хозяев. Ситуация изменяется в антропогенных экосистемах (особенно в сельскохозяйственных), где заражение паразитами может привести к существенному падежу скота.
Основная особенность отрицательного взаимодействия популяций заключается в том, что при их синхронной эволюции в стабильной экосистеме степень отрицательного влияния уменьшается. Иными словами, естественный отбор стремится уменьшить отрицательные влияния или вообще устранить взаимодействие популяций, поскольку продолжительное и сильное подавление популяции добычи или хозяина популяцией хищника или паразита может привести к уничтожению одной из них или обеих.
Материально - энергетический баланс производства. Потоки энергии в производственных процессах. Энергетическая эффективность деятельности. Функционально-стоимостной анализ как метод оптимизации.
Для описания технологических процессов широко используют материальные и энергетические балансы. Материальный баланс — это отражение закона сохранения массы веществ в условиях производства. Согласно этому закону, масса исходных веществ (сырья), поступивших для участия в технологическом процессе, равна массе веществ (продуктов и отходов), образовавшихся в результате осуществления технологического процесса. Как правило, материальный баланс составляется на единицу целевого продукта. Энергетический баланс — это количественное выражение закона сохранения энергии в ходе осуществления технологического процесса. Согласно этому закону, количества энергии, потребленной и выделяющейся в ходе процесса, равны. Как и материальный, энергетический баланс составляется на единицу целевого продукта. Материальные и энергетические балансы имеют большое значение для анализа и эффективного осуществления производственного процесса. С их помощью устанавливают фактический выход продукции, коэффициенты полезного использования энергии, расходы и потери сырья, топлива и других материалов. По цифровым данным технологических балансов принимаются организационные и технические решения по совершенствованию работы оборудования, максимальному использованию, утилизации или регенерации материальных и энергетических ресурсов. Балансы весьма важны для составления рациональных схем технологического процесса и установления оптимальных размеров, мощности и производительности оборудования. Данные технологических балансов и выводы из их анализа широко используются технологами и экономистами при проектировании новых и улучшении работы существующих предприятий.
ФУНКЦИОНАЛЬНО-СТОИМОСТНЫЙ АНАЛИЗ (ФСА) — это практический метод комплексного системного исследования объектов, направленный на обеспечение минимальных затрат на производство, реализацию или эксплуатацию при обеспечении необходимых потребительских свойств.
ФСА от других методов повышения эффективности производства отличает совокупность принципов, которая включает плановый характер проведения ФСА, комплексный подход, системный подход, функциональный подход, принципы соответствия значимости функций и затрат на их осуществление, соответствия фактического значения параметра требуемому уровню, активизации творческого мышления, коллективного труда, междисциплинарного подхода и др.
Сопряженная эволюция паразита и хозяина характеризуется совершенствованием механизмов противопаразитной борьбы у хозяина и повышение приспособленности паразитов.
У хозяина вырабатывается ряд защитных реакций, которые делятся на специфические под хозяина или паразита подстройка и неспецифические (изначально заложены, имунный ответ).
К неспецифическим относим иммунный ответ орг-ма, т. е. возникновение биохим реакций, кот сдерживают массу развития параз-в;
- сбрасывание заражен частей (это особенно характерно для Р-хоз, которые сбрасывают сильно зараженные листья). В этом случае паразиты продолжает жить уже как детритофаги;
- выработка устойчивости к вл параз за счет быстрого роста здоровых тканей взамен поражен (это имеет место при поедании тканей растений тлей);
- уменьшение плотности популяции хозяина, что снижение вероятности распространения паразитом и заражения им. Зараженные животные менее подвижно и становится более легкой добычей хищника, таким образом, снижает долю зараженной особей в популяции;
- формирование гетероген популяций хозяина, в составе которого есть экотипы, устойчивые к паразитам. Эти экотипы являются основой адаптивной селекции на повышение устойчивости культурных растений к грибковым заболеваниям.
У паразитов также в ходе совместной эволюции вырабатывается защитный механизм:
- инцистир и инкапсул, капсулы вокруг паразита формируются из тканей хозяина. Такая защитная изоляция возможна лишь у покоящихся стадий паразита, которая характеризуется минимальным уровнем обмена веществ.
- Локализм в тканях наиболее изолированных от других иммунных систем. Чтобы избежать атаки защит сих хоз, некоторые паразиты поселяются в наиболее защищенных от воспалит р-ий тканях. Н, метацеркарии некоторых трематод.
- Иммуносупрессия – способности подавления иммунной защиты хоз.
- Изменение состава антигенов.
- Молекулярная мимикрия – паразит покрывает себя веществами, образоваными хозяином, благодаря чему скрывается от распознания средствами его иммунной системы. К такой мимикр способн церкарии шистосом.
В процессе адаптации к своим хозяинам у паразитов происходит морфофизиол регресс органов, функции которого не востребованы. Наряду с этим у паразита развиваются физиологические механизмы и стр-рные приспособл, необх д сущ-я в орг-ме хоз.
- Изменяется форма тела. У эктопаразитов тело сплющивается в дорсо-вентрал направл, что облегч удержан на теле хоз. У блох же тело сплющен ос боков. Внутриполостные паразиты имеют компактное тело.
- Размеры обычно коррелируют с размерами соответств групп свободножив хоз. В некот случаях паразитизм приводит к увеличением размеров тела. Но размеры параз всегда много меньше размер хоз.
- Большим изменен подверг пищеварительная система, у некоторых она совсем отсут (ленточные черви). У кровосос она им большую емкость и способна вместить значит кол-во крови.
- Разнообразные приспособления для удержан в / на орг-ме хоз. Различают разнообразные крючки и присоски. Крючки встреч, нач с простейших (у грегарин) вплоть до членистон. Помимо органов присосок и крючков органами прикрепл служат различ стилеты, стрекат и липкие нити, стебельки и другие приспособл.
Вместе с морфол измен у параз происх и физиол адаптации к паразит обр ж
- Внутриполостн и ткан паразиты приобрели способность к анаэроб дых
- Кишечн паразиты приобрели способ вырабат антиферменты, кот защищ их от пищевар фермен хоз.
- Некот параз (лич шистосом и анкилостомид) стали выраб ферменты, с пом кот получили возм проникать через кож покровы в орг-м хоз.
- Большая плодовитость, гермафродитизм также явл приспособл к паразит обр ж.
Для естеств ЭС формир эко равновесия м-у популяциями параз и их хоз — норм явление. Во многих случаях проявл закономерн: плотность попул обоих видов измен циклич, но пики плотн параз запазд по отнош к пикам плотн хоз. Ситуация измен в антропог ЭС (особ в с/х), где зараж параз м привести к существенн падежу скота.
Осн особенность отрицат взаимод попул закл в т, чт при их синхронной эвол в стабил ЭС степень отрицат влияния уменьшается. Иными словами, естественный отбор стремится уменьшить отрицат вл / вообще устранить взаимод попул, поскольку продолжит и сильное подавл попул добычи / хоз попул хищника / паразита м привести к уничтож 1й из них / обеих.
7) АЛЛЕОПАТИЯ (АНТИБИОЗ). АМЕНСАЛИЗМ
Аллелопатия (антибиоз) — частный случай аменсализма, при котором во внешнюю среду выделяются продукты жизнедеятельности одного организма, отравляя ее и делая непригодной для жизни другого. Распространена у растений, грибов, бактерий. Сводится в конечном итоге к накоплению в корнеобитаемой среде определенных биологически активных химических соединений (чаще фенолов), в частности именно с помощью фенольных кислот растения бамбука подавляют рост и развитие проростков арахиса и кукурузы. Разлагающиеся листья каштана выделяют в почву токсичные соединения, подавляющие рост сеянцев деревьев многих других пород. Еще более интересен шалфей, листья которого продуцируют летучие терпены ( органические соединения, к которым относится камфора ), оказывающие воздействие на растущие поблизости растения непосредственно через атмосферу.
Аменсали́зм (от лат. - трапеза) - тип межвидовых взаимоотношений, при котором один вид, именуемый аменсалом, претерпевает угнетение роста и развития, а второй, именуемый ингибитором, таким испытаниям не подвержен.
Широко распространен во взаимоотношениях видов растений и ингибирующим фактором является либо отрицательное средообразование (одностороннее или, реже, взаимное), либо выделение ингибитором в окружающую среду каких-либо прижизненных выделений, отрицательно воздействующих на растение-аменсал (аллелопатия).
Примером одностороннего отрицательного средообразования может служить влияние деревьев-доминантов на виды мохового и травяного ярусов. Под пологом деревьев уменьшается освещённость, повышается влажность воздуха. При разложении опада деревьев почвы обедняются, поскольку при этом образуются кислоты, способствующие вымыванию элементов минерального питания вглубь почвенного слоя. Этот процесс особенно активен в таежном еловом лесу, так как ель — сильный средообразующий вид. Выносливые виды, участвующие в напочвенном покрове, компенсируют пагубность этого влияния и обеспечивают экологическое равновесие в таком лесу. При этом деревья (ингибиторы) не вступают в конкурентные отношения с видами напочвенного покрова (аменсалами), так как конкуренция подразумевает соревнование между видами при использовании определённого ресурса среды.
8) КОНКУРЕНЦИЯ И СОСУЩЕСТВОВАНИЕ ВИДОВ. ПРИНЦЫП ГАУЗЕ, ЭКСПЛУАТАЦИЯ И ИНТЕРФЕРОННАЯ КОНКУРЕНЦИЯ.
Конкуренция – это взаимоотношения видов со сходными экологическими требованиями существующих за счет общих ресурсов, имеющихся в недостатке. Когда такие виды обитают совместно, каждый из них находится в невыгодном положении, так как присутствие другого уменьшает возможности в овладении пищей, убежищами и прочими средствами к существованию, которыми располагает местообитание.
Конкуренция – единственная форма экологических отношений, отрицательно сказывающаяся на обоих взаимодействующих партнерах.
Формы конкурентного взаимодействия могут быть самыми различными: от прямой физической борьбы до мирного совместного существования.
Несовместимость конкурирующих видов еще раньше была подчеркнута Ч. Дарвином, который считал конкуренцию одной из важнейших составных частей борьбы за существование, играющей большую роль в эволюции видов.
Конкуренцией называется такое отношение между двумя популяциями, при котором увеличение каждой из них вызывает снижение численности другой. Впрочем, это определение касается только межвидовой конкуренции, а кроме нее существует также внутривидовая конкуренция, которая разворачивается внутри одной популяции.
При конкуренции две популяции могут неблагоприятно влиять друг на друга двумя разными способами. Первый (косвенная, или эксплуатационная конкуренция) связан с тем, что две популяции используют один и тот же ресурс. При возрастании численности одной из этих популяций усилится потребление ею общего ресурса, и другой достанется меньшее его количество. Второй (прямая, или интерференционная конкуренция) связан с затратами энергии особей на причинение друг другу ущерба. Примером прямой конкуренции может быть аллелопатия — выделение растениями веществ, угнетающих другие виды.
Полное замещение одного вида другим — не единственный результат межвидовой конкуренции, о чем свидетельствует часто наблюдаемое в природе сосуществование близких видов со сходными экологическими потребностями. Существует ряд ситуаций, в которых полное замещение невозможно. Четыре из них приведены ниже.
1. Замещение одного вида другим — длительный процесс. Поэтому следует ожидать, что, производя наблюдения в любой данный момент времени, мы обнаружим несколько пар конкурирующих видов, находящихся на стадии незавершенного замещения.
По существующим оценкам число фитопланктонных видов в некоторых озерах больше, чем число ниш для такого планктона. Это превышающее ожидания разнообразие видов объясняется, возможно, тем, что межвидовая конкуренция ещё не завершилась.
2. Экологически сходные виды могут сосуществовать, никогда не достигая стадии прямой межвидовой конкуренции. Так будет обстоять дело в том случае, если численность этих видов сдерживается каким-то другим фактором, а не прямой конкуренцией.
Численность популяций растительноядных животных в природных сообществах нередко удерживается на низком уровне хищниками и, следовательно, хищничество служит препятствием к тому, чтобы межвидовая конкуренция за пищу между растительноядными превратилась в важный фактор, оказывающий влияние на сосуществование этих последних.
3. Условия среды могут обратимо изменяться в период действия межвидового отбора, в результате чего на одной стадии селективным преимуществом обладает вид А, а на другой — вид В. В таком случае эти два вида будут сосуществовать в условиях циклического равновесия.
В некоторых временных водоемах обитают совместно разные зеленые водоросли, относящиеся к родам Haematococcus, Chlamydomonas, Scenedesmus и Chlorella. Для представителей Haematococcus благоприятны условия, создающиеся при пересыхании водоема,а для остальных водорослей — стоячая вода. При частой смене пересыхания и наполнения водоема сосуществование этих противоположных по своим предпочтениям зеленых водорослей может продолжаться бесконечно долго.
4. Очень важное значение в природных условиях имеет ситуация, при которой окружающая среда неоднородна, различаясь в разных своих участках по некоторому критическому фактору. При этом вид А может превосходить вид В в одном участке среды, а вид В — обладать преимуществом в другом её участке; в такой ситуации виды А и В могут сосуществовать, обитая частично или главным образом в предпочитаемом каждым из них участке среды.
В этом отношении весьма показательны лабораторные эксперименты с Paramecium. Следует напомнить, что P. aurelia замещает P. caudatum или наоборот в гомогенной культуральной среде (Гаузе, 1934*). Если, однако, лабораторная среда гетерогенна, то замещения может и не произойти. Смеси P. aurelia и P. bursaria помещали в пробирки, содержавшие вертикально стратифицированные суспензии кормовых дрожжей. В этом случае P. aurelia питается главным образом в верхних слоях, а P. bursaria — на дне. При создании таких разных кормовых ниш и вследствие различных пищевых предпочтений эти два вида могут сосуществовать неопределённо долгое время .
Естественные местообитания, конечно, всегда неоднородны, причём часто такая неоднородность допускает некоторую экологическую сегрегацию симпатрических видов, а следовательно, непрерывное сосуществование.
9) Роль конкуренции в разделении местообитаний
При наличии конкурентні с близкородственными или экологически сходными видами диапазон местообитаний, возможных для данного вида, сокращается до оптимальных границ (т. е. вид распространяется в наиболее благоприятных для него местообитаниях, в которых он обладает какими-либо преимуществами по сравнению со своими конкурентами). Если же межвидовая конкуренция выражена слабо, то под Влияние конкуренции на распространение птиц в местообитаниях. Щели преобладает внутривидовая конкуренция, то вид широко распространяется, занимая и менее благоприятные (краевые) области. Если же ин^ тенсивна межвидовая конкуренция, то распространение ограничивается •узкими пределами, где имеются оптимальные условия. Влиянием внутривидовой конкуренции популяции данного вида расширяют границы своего местообитания.
На островах, которые не заселены потенциальными конкурентами, особенно хорошо прослеживается тенденция к расширению местообитаний. Например, на островах, где отсутствуют лесные шолевки (Clethrionomys), луговые серые полевки (Microtus) часто занимают лесные местообитания (см. Cameron, 1964). Кроуэлл (Crowell, 1962) обнаружил, что на Бермудских островах кардиналы более многочисленны и занимают более краевые по условиям местообитания, так как здесь нет многих из его материковых конкурентов.
Хорошим примером экологического разделения сфер питания смогут служить две родственные рыбоядные птицы Великобритании, большой баклан (Phalacrocorax carbo) и длинноносый, или хохлатый, баклан (P. aristotelis), исследованные Лэком (Lack, 1945). Эти два вида обычно кормятся в одних и тех же водах ги гнездятся на одних и тех же обрывах, однако более глубокий анализ показывает, что на самом деле места их гнездовий различаются, а, кроме того, существенные различия выявляются и в составе их пищи (рис. 7.4): длинноносый баклан ловит рыбу, плавающую в верхних слоях воды, тогда как большой баклан добывает пищу в основном у дна, где он ловит камбал и донных -беспозвоночных (креветок и др.).
Тот факт, что близкородственные виды в природе четкэ разграничены, вовсе не означает, что для поддержания этой разобщенности между ними непрерывно должна поддерживаться конкуренция. В процессе эволюции у этих видов могут выработаться различные потребности и предпочтения, и тогда конкурентные отношения между ними прекращаются.
10) КОНЦЕПЦИИ МЕСТООБИТАНИЯ, ЭКОЛОГИЧЕСКОЙ НИШИ И ГИЛЬДИИ
Местообитание организма - это место, где он живет, или место, где его обычно можно найти.
Экологическая ниша - понятие более емкое, включающее в себя не только физическое пространство, занимаемое организмом, но и функциональную роль организма в сообществе (Н, его трофическое положение) и его положение относительно градиентов внешних факторов — температуры, влажности, рН, почвы и других условий существования. Эти три аспекта экологической ниши удобно обозначить как пространственную нишу, или нишу места, трофическую пишу и многомерную нишу, или нишу как гиперобъем. Следовательно, экологическая ниша организма зависит не только от того, где он живет, но включает также общую сумму его требований к окружающей среде. Концепция ниши оказывается наиболее полезной для количественных оценок различий между видами (или внутри вида в разных местах и в разное время) по одному или немногим основным (т. е. операционально значимым) признакам. Для характеристики ниши чаще всего используются два измерения — ширина ниши и перекрывание ниши с соседними. Группы видов в сообществе, обладающих сходными функциями и нишами одинакового размера, называются гильдиями. Виды, занимающие одинаковые ниши в разных географических областях, называются экологическими эквивалентами.
Местообитание водных насекомых: клопа-гладыша и плавта - это мелководные, покрытые растительностью пространства (литоральная зона) прудов и озер. Эти виды занимают, однако, очень разные трофические ниши. Гладыш - активный хищник, а плавт питается преимущественно разлагающейся растительностью.
Термин гильдия часто используется в применении к группам видов роль которых в сообществе одинакова или сравнима Н. осы, паразитирующие на популяциях питающихся нектаром насекомых, улитки, живущие в листовой подстилке леса, и лианы, проникающие в полог тропического леса.
Гильдия — удобная единица для изучения взаимодействий между видами, однако при анализе сообщества ее можно рассматривать как функциональную единицу, в результате чего отпадает необходимость рассматривать все до единого виды по отдельности.
Местообитание – это территория или акватория, занимаемая популяцией (видом), с комплексом присущих ей экологических факторов. Местообитание вида является компонентом его экологической ниши. Применительно к наземным животным местообитание вида называется стация. Местообитание сообщества – биотоп.
Для характеристики экологической ниши обычно используют два важных показателя: ширина ниши и степень перекрывания ее с соседними. Экологические ниши разных видов могут быть разной ширины и перекрываться в различной степени.
Несовместимость конкурирующих видов еще раньше была подчеркнута Ч. Дарвином, который считал конкуренцию одной из важнейших составных частей борьбы за существование, играющей большую роль в эволюции видов.
В опытах Г. Ф. Гаузе с культурами инфузорий Paramecium aurelia и Р. caudatum каждый из видов, помещенных отдельно в пробирки с сенным настоем, успешно размножался, достигая определенного уровня численности. Если же оба вида со сходным характером питания помещали совместно, то первое время наблюдался рост численности каждого из них, но затем количество Р. caudatum постепенно сокращалось, и они исчезали из настоя, тогда как количество Р. aurelia оставалось постоянным.
Победителем в конкурентной борьбе оказывается, как правило, тот вид, который в данной экологической обстановке имеет хотя бы небольшие преимущества перед другим, т.е. больше приспособлен к условиям окружающей среды, поскольку даже близкие виды никогда не совпадают по всему экологическому спектру.
Экологические ниши видов изменчивы в пространстве и во времени. Они могут быть резко разграничены в индивидуальном развитии в зависимости от стадии онтогенеза, как, например у гусениц и имаго чешуекрылых, личинок и жуков майского хруща, головастиков и взрослых лягушек. В этом случае меняется и среда обитания, и все биоценотическое окружение.
Группы видов в сообществе, обладающие сходными функциями и нишами одинакового размера, то есть роль которых в сообществе одинакова или сравнима, называются гильдиями. Например, лианы тропического леса представлены многими видами растений. Между видами внутри гильдии наблюдается особенно острая конкуренция.
Виды, занимающие одинаковые ниши в разных географических областях, называются экологическими эквивалентами. Например, крупные кенгуру Австралии, бизоны Северной Америки, зебры и антилопы Африки и т. д. являются экологическими эквивалентами. В настоящее время они значительно замещены коровами и овцами.
Правило географического оптимума – в центре видового ареала, как правило, имеются оптимальные для вида условия существования, которые ухудшаются к периферии области его обитания.
Не нашли то, что искали? Воспользуйтесь поиском:
Читайте также:
