
Что такое цикл кори
ОБМЕН УГЛЕВОДОВ
Классификация:
1) Моносахариды – глюкоза, фруктоза, галактоза.
2) Олигосахариды – мальтоза, сахароза, лактоза.
3) Полисахариды – крахмал и гликоген.
Гликоген – полисахарид в котором молекулы глюкозы соединены 1,4-гликозидными связями, а в местах ветвления 1,6-гликозидными связями.
Крахмал – гетерополисахарид, состоящий из линейной амилозы и разветвленного амилопектина. Строительная единица крахмала – глюкоза соединенная 1,4- и 1,6-гликозидными связями.
Переваривание углеводов:
Начинается в ротовой полости, под действием амилазы слюны, которая является эндоамилазой, то есть разрывает внутренные 1,4-гликозидные связи. Действие про pH=6,8-7.0
Амилаза в активном центре содержит Ca, активатором амилазы является анион Cl - .
Глубокого переваривания в ротовой полости не происходит, так как пища там находится недолго. Образуются промежуточные продукты гидролиза крахмала – декстрины(амило-, эритро-, мальто-, ахродекстрины).
При попадании пищевого комка в желудок амилаза слюны инактивируется, так как pH желудочного сока 1,5-2,5
Переваривание углеводов внутри пищевого комка не происходит, так как соляная кислота не попадает внутрь пищевого комка. Основное переваривание углеводов происходит в кишке под действием панкреатической амилазы(диастаза), которая разрывает внутренние 1,4-гликозидные связи.
Панкреатический сок содержит еще 2 фермента – Амило- и олиго-1,6-гликозидазы, которые разрывают 1,6-гликозидные связи.Конечными продуктами переваривания крахмала являются мальтоза, мальтотриоза, глюкоза.
Кишечный сок содержит сахаразу, которая действует на сахарозу и расщепляет её на сахарозу и фруктозу. Так же в нем есть мальтаза, которая действует на мальтозу, расщепляя ее на 2 остатка глюкозы. Лактаза действует на лактозу и расщепляет ее на глюкозу и галактозу.
Конечные продукты переваривания полисахаридов являются моносахариды. Образовавшиеся моносахариды всасываются с различной скоростью, скорость всасывания глюкозы 100 процентов.
Использование глюкозы .
1) синтез гликогена в печени и скелетных мышцах. Наибольшее количество гликогена откладывается в мышцах, запас гликогена хватает на 24 часа. Гликоген откладывается в виде гранул или глыбок, где содержаться ферменты и синтеза и распада гликогена.
Глюкоза, попав в клетку активируется или фосфорилируется с образованием глюкозо-6-фосфата. Эту реакции катализирует ферменты – глюкокиназа или гексокиназа, которая обладает различным сродством к глюкозе. Реакция не обратимая и требует АТФ. Глюкозо-6-фосфат основной метаболит обмена углеводов.
2) Глюкозо-6-фосфат(фермент изомераза)→ глюкоза-1-фосфат.
3) Глюкоза-1-фосфат взаимодействует с УТФ в результате образуется транспортная форма глюкозы при синтезе гликогена УДФ-глюкоза. Фермент реакции трансфераза.
4) УДФ-глюкоза передает свой остаток глюкоза-1-фосфат на затравочное количество гликогена с образование 1,4-гликозидной связи. Фермент гликогенсинтетаза – главный фермент синтеза гликогена, может существовать в двух формах: фосфорилированная неактивная, дефосфорилированная активная. В образовании 1,6-гликозидных связей участвует фермент ветвления.
Распад Гликогена
Протекает при участии фермента гликоген-фосфарилазы, который может находится в 2 формах:
Активации гликоген-фосфорилазы осуществляют гормоны адреналин и глюкагон.
Адреналин действует при стрессах , активирует распад гликогенав печени и скелетных мышцах. Глюкагон действует в норме и в постабсортивный период, активируя распад гликогена только в печени.
Механизм действия глюкагона и адреналина
Так как для этих гормонов мембрана клетки не проницаема, рецепторы для них располагается на внешней поверхности клеточной мембраны. Происходит образование гормон-рецепторного комплекса, затем изменяется конформация G-белка за счет гидролиза ГТФ в ГДФ. G-белок расположен в самой мембране.
Активируется аденилатциклаза, расположенная на внутренней поверхности мембраны. Активированная аденилатциклаза в клетке катализирует реакцию образования цАМФ из АТФ.
цАМФ вторичный посредник или мессенджер в передаче гормонального сигнала внутрь клетки. На этом этапе происходит усиление гормонального сигнала цАМФ образуется много до 500 молекул, но время их жизни очень мало! так как они быстро гидролизируются ферментами. цАМФ активирует протеинкиназу, актиная протеинкиназа фосфорилирует белки-ферменты клетки. Активная гликоген-фосфорилаза отщепляет от гликогена глюкоза-1-фосфат→глюкозо-6-фосфат→глюкоза→в кровь.
Гликолиз
Протекает в цитоплазме может быть аэробный и анаэробный.
Анаэробный гликолиз – это окисление глюкозы в отсутствии кислорода до двух молекул лактата и 2-х АТФ.
1) Образование глюкозо-6-фосфат. Реакция необратима, затрачивается 1 молекула АТФ.
3) Фруктозо-6-фосфат при участии АТФ и фермента фосфофруктокиназы фосфорилируется с образованием фруктоза-1,6—дифосфат. Реакция необратима, затрачивается АТФ. Эта реакция определяет скорость гликолиза в целом. Фосфофруктокиназа алостерический фермент его активность угнетается высоким содержанием АТФ и повышается при высоких концентрациях АМФ в клетке.
4) Фруктоза-1,6-фосфат при участии фермента альдолаза распадается на 2 фосфотриозы: диоксиацетонфосфат, глицероальдегидтрифосфат.
На этом заканчивается неокислительная стадия гликолиза и начинается стадия гликолитической оксиредукции, в которую вступают 2 молекулы глицероальдегиттрифосфат.
6) Реакция окислительного фосфорилирования протекает при участии неорганического фосфата. (H3PO4) НАД и фермента глицероальдегидфосфатдегидрогиназа. В результате образуется 1,3-дифосфоглицерат + 2НАДН2.
7) Субстратное фосфорилирование. 1,3-дифосфоглицерат передает свой остаток фосфата из первого положения на АДФ в результате образуется 2 молекулы АТФ и 2 молекул 3-фосфоглицерата. Фермент фосфоглицераткиназа.
9) 2-фосфоглицерат(енолаза)→фосфоенолпируват содержит макроэргическую связь, обозначаемую значком
10) Субстратное фосфорилирование. 2-фосфоенолпируват + 2АДФ→2пируват + 2АТФ. Фермент пируваткиназа, реакция необратима.
11) При участии НАДН2, которые образовались в 6-й реакции, восстанавливается в лактат. Фермент лактат дегидрогиназа.
Энергетический выход анаэробного гликолиза:
1-3-й реакции затрачивается 2 молекулы АТФ. 7,10-й реакции образуется 4 молекулы АТФ. И того 4-2=2 молекулы АТФ.
Биологическое значение анаэробного гликолиза.
Заключается в том что клетка получает 2 молекулы АТФ в отсутствии кислорода.
Аэробный гликолиз – это окисление глюкозы в присутствии кислорода с образованием 2 молекул пирувата и 7 молекул АТФ. Аэробный гликолиз состоит из 10 реакций. Энергетический выход аэробного гликолиза:
1-3-й реакции затрачивается 2 молекулы АТФ. 6-й реакции 2 молекулы НАДН2, которые поступают в дыхательную цепь и там дают 2.5 молекулы АТФ*2=5 молекул АТФ.(одна молекула НАДН2 дает 2.5 молекул АТФ, а одна молекула ФАДН2 дает 1.5 молекулы АТФ). 7,10-й реакции образуется 4 молекулы АТФ. Итого 5+4-2=7 молекул АТФ.
Анаэробный гликолиз отличается от аэробного гликолиза:
1) условие проведения
2) количество реакций анаэробный – 11, аэробный – 10.
3) Конечные продукты анаэробный – 2 молекулы лактата, аэробный – 2 молекулы пирувата.
4) Количество энергии анаэробный – 2 молекулы АТФ, аэробный – 7 молекул АТФ.
Глюконеогенез
Это синтез глюкозы из неуглеводных продуктов – пируват, лактат, гликогенные аминокислоты(белки), глицерин(жиры). Синтезировать глюкозу из пирувата и лактата по пути гликолиза невозможно, так как 3 реакции гликолиза являются необратимыми 1,3,10. Поэтому в глюконеогенезе обходные пути и специальные ферменты. Например, надо синтезировать глюкозу из 2-х молекул пирувата. Пируват не может превратится в фосфоенолпируват по пути гликолиза, так как 10-я реакция необратима.
1-я обходная реакция пируват в митохондриях при участии 2-х молекул АТФ и CO2 карбоксилируется в оксалоацетат. Фермент этой реакции пируваткарбоксилаза, Кофермент витамин Н(биотин), при нехватки витамина активность фермента снижается. Оксалоацетат не может попасть в цитозоль, так как мембрана митохондрий для него не проницаема, сначала оксалоацетат превращается в малат, который переходит в цитозоль и вновь превращается в оксалоацетат. Оксалоацетат при участии 2-х молекул ГТФ и фермента фосфоенолпируваткарбоксикиназа превращается в фосфоенолпируват. Далее идут реакции гликолиза фосфоенолпируват→2-фосфоглицерат→3-фосфоглицерат(2АТФ) →1,3-дифосфоглицерат→2 молекулы глицероальдегидтрифосфат (1 молекула дает фрутозо-1,6-дифосфат) другая молекула дает диоксиацетонфосфат→фрутозо-1,6-дифосфат, который не может превратится во фрутоза-6-фосфат по пути гликолиза, так как 3-я реакция гликолиза необратима.
2-я обходная реакция ее катализирует фермент фруктозодифосфотаза при участии которого фрутозо-1,6-дифосфат превращается во фруктозо-6-фосфат→глюкозо-6-фосфат, который не может превратится в свободную глюкозу по пути гликолиза, так как 1-я реакция гликолиза необратима.
3-я обходная реакция катализирует фермент глюкозо-6-фосфатаза. На синтез 1 молекулы глюкозы из 2-х молекул пирувата тратится 6 молекул АТФ. Глюконеогенез контролтрует кортизол.
Глюкозолактатный цикл(цикл Кори).
Гликолиз, который протекает в скелетных мышцах связан с глюконеогенезом, который протекает в печени. При мышечной работе в них накапливается лактат, который выходит в кровь и попадает в печень, где из него синтезируется глюкоза путем глюконеогенеза. Эта глюкоза выходит в кровь, затем попадает в мышцы где используется в качестве энергетического материала.
Пировиноградная кислота, которая образовалась в результате аэробного гликолиза, подвергается окислительному карбоксилированию с образованием ацетилКоА и НАДН2(2.5 АТФ). Эту реакцию катализирует пируватдегидрогиназный комплекс, который состоит из 3 ферментов и 5 коферментов.
5 коферментов: Тиаминпирофосфат, который связан с пируватдекарбоксилазой , липоат(липоевая кислота), Коэнзим А, ФАД, НАД.
[youtube.player]
Цикл Кори – это важный метаболический процесс, который помогает нашему организму вырабатывать дополнительное количество энергии, требующееся мышцам для осуществления физической деятельности повышенной интенсивности. Чтобы понять, как работает организм человека, важно проанализировать многочисленные менее масштабные процессы, происходящие внутри него. Вместе эти автономные и неавтономные процессы, следуя один за другим, позволяют нам жить и заниматься нашими повседневными делами.
Выработка энергии для мышечной активности
Мышцы в нашем организме позволяют нам осуществлять все виды нашей повседневной деятельности, включая хождение, стояние, бег, поднятие тяжестей, и т.д. Они обеспечивают количество силы прямо пропорциональное интенсивности осуществляемой деятельности. Для генерирования этой силы необходима энергия. Мышечная активность требует аденозинтрифосфата (АТФ), который вырабатывается в ходе процесса известного как гликогенолиз. Гликогенолиз – это распад с высвобождением глюкозы гликогена, который хранится в скелетных мышцах.
При осуществлении большинства видов нашей повседневной деятельности наши мышцы совмещают глюкозу и кислород аэробно, в ходе процесса известного как гликолиз, который приводит к выработке двух молекул АТФ и двух молекул пирувата.
АТФ используется для выработки энергии непосредственно. Пируват распадается далее, когда доступно достаточно кислорода, генерируя еще большее количество энергии. Таким образом, эти две метаболические составляющие обеспечивают мышцы энергией на клеточном уровне, давая им возможность функционировать.
Тем не менее, когда мы осуществляем физическую деятельность повышенной интенсивности, количество поступающего в организм кислорода становится диспропорциональным (гораздо меньшим) относительно потребности мышц в энергии. При таком сценарии из-за того, что кислорода недостаточно, глюкоза расщепляется посредством анаэробного метаболического процесса, известного как ферментация, в ходе которого пируват преобразуется в лактат – растворимую молочную кислоту, а затем выделяется в кровь.
Это позволяет химическим процессам, ответственным за выработку энергии, продолжатся без применения кислорода. Мышечные клетки могут вырабатывать энергию анаэробно таким способом на очень высокой скорости, но только в течение одной-трех минут, после чего количество лактата в крови становится избыточным, что приводит к усталости.
Что такое цикл Кори?
Если интенсивная деятельность продолжается, организм прибегает к альтернативному метаболическому пути для устранения лактата и продолжения выработки энергии анаэробно. Этот процесс выработки энергии называется циклом Кори.
Цикл Кори
В ходе работы цикла Кори лактат, скапливающийся в мышечных клетках забирается печенью. Печень осуществляет химический процесс, известный как глюконеогенез, представляющий собой преобразование лактата обратно в глюкозу. По сути, глюконеогенез дает обратный ход и процессу гликолиза, и процессу ферментации, которые осуществляются организмом для выработки лактата. Он сначала преобразует лактат в пируват, а затем, наконец, в глюкозу.
Эта глюкоза впоследствии попадает в кровоток, который доставляет ее к работающим мышцам, где она используется для удовлетворения потребности мышц в дополнительной энергии. Последующая выработка мышцами лактата опять нейтрализуется печенью, таким образом, цикл Кори возобновляется. В случае если мышечная активность прекращается, глюкоза, вырабатываемая в ходе работы цикла Кори, подвергается гликогенезу и используется для пополнения запасов гликогена в мышцах.
Ограниченность цикла Кори
Используя цикл Кори, организм способен преобразовывать побочные продукты метаболизма в источник энергии для мышц. Однако он не может делать это бесконечно. Так как же многие другие естественные циклы, цикл Кори не является полностью замкнутым циклом. Гликолиз приводит к выработке в мышцах двух молекул АТФ. Однако печень использует для осуществления процесса глюконеогенеза до шести молекул АТФ. Цикл Кори также требует первоначального поступления кислорода, без которого он не может начинаться. По этой причине, мышцам, в конечном итоге, неизбежно требуется новый запас глюкозы, а также кислорода.
Если физическая деятельность становится слишком интенсивной, потребности мышц в энергии превосходят возможности цикла Кори по регенерированию глюкозы из лактата.
Это приводит к состоянию, известному как лактатацидоз, который представляет собой чрезмерное накопление молочной кислоты в организме. Лактатацидоз понижает уровень pH крови, что может вызывать повреждение тканей. Он также провоцирует появление симптомов, связанных с паникой, таких как гипервентиляция, спазмы в животе, рвота и т.д. – все это природные защитные механизмы организма, направленные на прекращение интенсивной деятельности и предупреждение возникновения перманентных повреждений.
[youtube.player]Некоторые ткани, например, мозг, нуждаются в постоянном поступлении глюкозы. Когда поступление углеводов в составе пищи недостаточно, содержание глюкозы в крови некоторое время поддерживается в пределах нормы за счёт расщепления гликогена в печени. Однако запасы гликогена в печени невелики. Они значительно уменьшаются к 6 —10 ч голодания и практически полностью исчерпываются после суточного голодания. В этом случае в печени начинается синтез глюкозы de novo — глюконеогенез.Глюконеогенез — процесс синтеза глюкозы из веществ неуглеводной природы. Его основной функцией является поддержание уровня глюкозы в крови в период длительного голодания и интенсивных физических нагрузок. Процесс протекает в основном в печени и менее интенсивно в корковом веществе почек, а также в слизистой оболочке кишечника. Эти ткани могут обеспечивать синтез 80 — 100 г глюкозы в сутки. На долю мозга при голодании приходится большая часть потребности организма в глюкозе. Это объясняется тем, что клетки мозга не способны, в отличие от других тканей, обеспечивать потребности в энергии за счёт окисления жирных кислот (см. раздел 8).
Кроме мозга, в глюкозе нуждаются ткани и клетки, в которых аэробный путь распада невозможен или ограничен, например, эритроциты (они лишены митохондрий), клетки сетчатки, мозгового слоя надпочечников и др.
Первичные субстраты глюконеогенеза — лактат, аминокислоты и глицерол. Включение этих субстратов в глюконеогенез зависит от физиологического состояния организма.
• Лактат — продукт анаэробного гликолиза. Он образуется при любых состояниях организма в эритроцитах и работающих мышцах. Таким образом, лактат используется в глюконеогенезе постоянно.
• Глицерол высвобождается при гидролизе жиров в жировой ткани в период голодания или при длительной физической нагрузке.
• Аминокислоты образуются в результате распада мышечных белков и включаются в глюконеогенез при длительном голодании или продолжительной мышечной работе.
На рисунке 7-44 показаны пункты включения первичных субстратов в глюконеогенез.
Рис. 7-44. Включение субстратов в глюконеогенез.
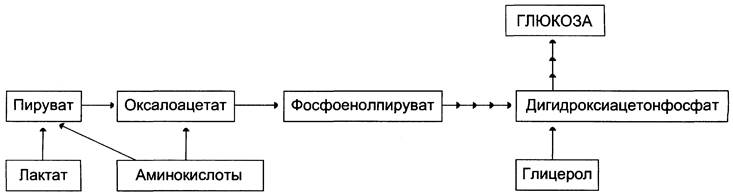
А. Реакции глюконеогенеза
Большинство реакций глюконеогенеза протекает за счёт обратимых реакций гликолиза (рис. 7-45, реакции 9, 8, 7, 6, 5, 4, 2) и катализируется теми же ферментами. Однако 3 реакции гликолиза термодинамически необратимы. На этих стадиях реакции глюконеогенеза протекают другими путями.
Рис. 7-45. Гликолиз и глюконеогенез. Ферменты обратимых реакций гликолиза и глюконеогенеза: 2 — фосфоглюкоизомераза; 4 — альдолаза; 5 — триозофосфатизомераза; 6 — глицеральдегидфосфатдегидрогеназа; 7 — фосфоглицераткиназа; 8 — фосфоглицератмутаза; 9 — енолаза. Ферменты необратимых реакций глюконеогенеза: 11 — пируваткарбоксилаза; 12 — фосфоенолпируваткарбоксикиназа; 13 — фруктозо- 1,6-бисфосфатаза; 14 — глюкозо-6-фосфатаза. I-III —субстратные циклы.
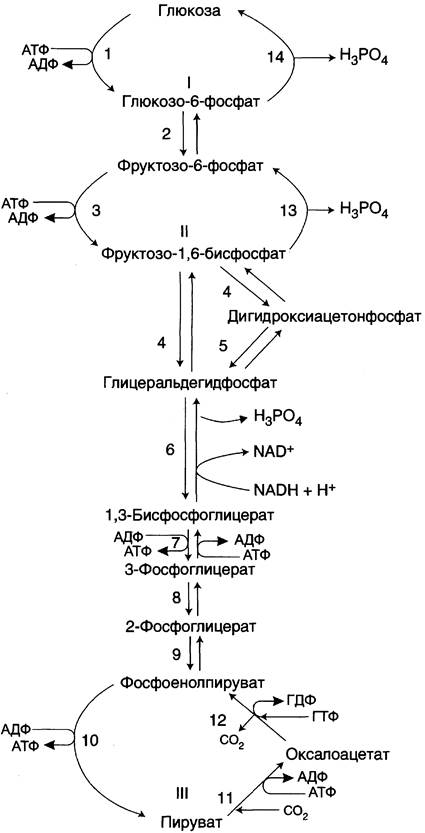
Необходимо отметить, что гликолиз протекает в цитозоле, а часть реакций глюконеогенеза происходит в митохондриях.
Рассмотрим более подробно те реакции глюконеогенеза, которые отличаются от реакций гликолиза и происходят в глюконеогенезе с использованием других ферментов. Рассмотрим процесс синтеза глюкозы из пирувата.
1. Образование фосфоенолпирувата из пирувата — первая из необратимых стадий глюконеогенеза
Образование фосфоенолпирувата из пирувата происходит в ходе двух реакций (рис. 7-45, реакции 11, 12), первая из которых протекает в митохондриях. Пируват, образующийся из лактата или из некоторых аминокислот, транспортируется в матрикс митохондрий и там карбоксилируется с образованием оксалоацетата (рис. 7-46). Пируват- карбоксилаза,катализирующая данную реакцию, — митохондриальный фермент, коферментом которого является биотин. Реакция протекает с использованием АТФ.
Рис. 7-46. Образование оксалоацетата из пирувата.
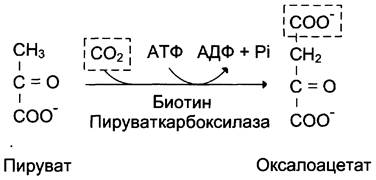
Дальнейшие превращения оксалоацетата протекают в цитозоле. Следовательно, на этом этапе должна существовать система транспорта оксалоацетата через митохондриальную мембрану, которая для него непроницаема. Оксалоацетат в митохондриальном матриксе восстанавливается с образованием малата (рис. 7-47) при участии НАDН (обратная реакция цитратного цикла). Образовавшийся малат затем проходит через митохондриальную мембрану с помощью специальных переносчиков. Кроме того, оксалоацетат способен транспортироваться из митохондрий в цитозоль в виде аспартата в ходе малат-аспартатного челночного механизма, рассмотренного ранее (рис. 7-39).
Рис. 7-47. Превращение оксалоацетата в малат.
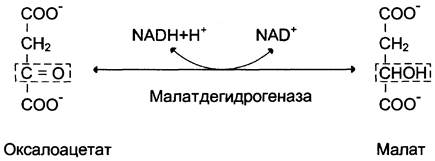
В цитозоле малат вновь превращается в оксалоацетат в ходе реакции окисления с участием кофермента NAD + . Обе реакции: восстановление оксалоацетата и окисление малата катализируют малатдегидрогеназа, но в первом случае это митохондриальный фермент, а во втором — цитозольный. Образованный в цитозоле из малата оксалоацетат затем превращается в фосфоенолпируват в ходе реакции, катализируемой фосфоенолпируваткарбоксикиназой — ГТФ-зависимым ферментом (рис. 7-48). Название фермента дано по обратной реакции.
Рис. 7-48. Превращение оксалоацетата в фосфоенолпируват.
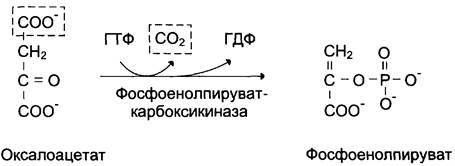
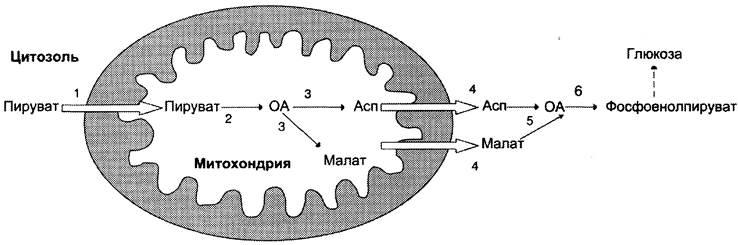
Схема всех реакций, протекающих на первой необратимой стадии глюконеогенеза, представлена на рис. 7-49.
Рис. 7-49. Образование оксалоацетата, транспорт в цитозоль и превращение в фосфоенолпируват. 1 — транспорт пирувата из цитозоля в митохондрию; 2 — превращение пирувата в оксалоацетат (ОА); 3 — превращение ОА в малат или аспартат; 4 — транспорт аспартата и маната из митохондрии в цитозоль; 5 — превращение аспартата и маната в ОА; 6 — превращение ОА в фосфоенолпируват.
Следует отметить, что этот обходной участок глюконеогенеза требует расхода двух молекул с макроэргическими связями (АТФ и ГТФ) в расчёте на одну молекулу исходного вещества — пирувата. В пересчёте на синтез одной молекулы глюкозы из двух молекул пирувата расход составляет 2 моль АТФ и 2 моль ГТФ или 4 моль АТФ (для удобства рассуждений предлагается считать, что энергозатраты на синтез АТФ и ГТФ равны). После образования фосфоенолпирувата все остальные реакции также
протекают в цитозоле вплоть до образования фруктозо-1,6-бисфосфата и катализируются гликолитическими ферментами.
2. Гидролиз фруктозо-1,6-бисфосфата и глюкозо-6-фосфата
Отщепление фосфатной группы из фруктозо-1,6-бисфосфата и глюкозо-6-фосфата — также необратимые реакции глюконеогенеза. В ходе гликолиза эти реакции катализируют специфические киназы с использованием энергии АТФ. В глюконеогенезе они протекают без участия АТФ и АДФ и ускоряются не киназами, а фосфатазами — ферментами, принадлежащими к классу гидролаз. Ферменты фруктозо-1,6-бисфосфатаза и глюкозо-6-фосфатаза катализируют отщепление фосфатной группы от фруктозо-1,6-бисфосфата и глюкозо-6-фосфата. После чего свободная глюкоза выходит из клетки в кровь. Схема всех реакций глюконеогенеза представлена на рис. 7-45.
Итак, в печени существуют 4 фермента, которые принимают участие только в глюконеогенезе и катализируют обходные реакции необратимых стадий гликолиза. Это — пируват- карбоксилаза, фосфоенолпируваткарбоксикиназа, фруктозо-1,6-бисфосфатаза и глюкозо-6- фосфатаза.
3. Энергетический баланс глюконеогенеза из пирувата
В ходе этого процесса расходуются 6 моль АТФ на синтез 1 моль глюкозы из 2 моль пирувата. Четыре моль АТФ расходуются на стадии синтеза фосфоенолпирувата из оксалоацетата и ещё 2 моль АТФ на стадиях образования 1,3-бисфосфоглицерата из 3-фосфоглицерата.
Суммарный результат глюконеогенеза из пирувата выражается следующим уравнением:
2 Пируват + 4 АТФ + 2 ГТФ + 2 (NАDН + Н + ) + 4 Н2O —> Глюкоза + 4 АДФ + 2 ГДФ + 6 Н3РO4 + 2 NАD + .
Б. Синтез глюкозы из лактата
Лактат, образованный в анаэробном гликолизе, не является конечным продуктом метаболизма. Использование лактата связано с его превращением в печени в пируват. Лактат как источник пирувата важен не столько при голодании, сколько при нормальной жизнедеятельности организма. Его превращение в пируват и дальнейшее использование последнего являются способом утилизации лактата.
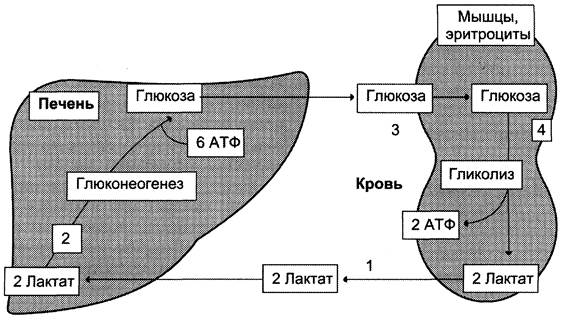
Лактат, образовавшийся в интенсивно работающих мышцах или в клетках с преобладающим анаэробным способом катаболизма глюкозы, поступает в кровь, а затем в печень. В печени отношение NАDН/NАD + ниже, чем в сокращающейся мышце, поэтому лакгатдегидрогеназная реакция протекает в обратном направлении, т. е. в сторону образования пирувата из лактата. Далее пируват включается в глюконеогенез, а образовавшаяся глюкоза поступает в кровь и поглощается скелетными мышцами. Эту
Рис. 7-50. Цикл Кори (глюкозо-лактатный цикл). 1 — поступление лактата из сокращающейся мышцы с током крови в печень; 2 — синтез глюкозы из лактата в печени; 3 — поступление глюкозы из печени с током крови в работающую мышцу; 4 — использование глюкозы как энергетического субстрата сокращающейся мышцей и образование лактата.
тиляции и, следовательно, ускорением выведения СO2 из организма. Однако основную роль в компенсации ацидоза играют почечные механизмы с участием аммиачного буфера (см. раздел 9). Одной из причин метаболического ацидоза может быть накопление молочной кислоты. В норме лактат в печени превращается обратно в глюкозу путём глюконеогенеза либо окисляется. Кроме печени, другим потребителем лактата служат почки и сердечная мышца, где лактат может окисляться до СO2 и Н2O и использоваться как источник энергии, особенно при физической работе.
Уровень лактата в крови — результат равновесия между процессами его образования и утилизации. Кратковременный компенсированный лактоацидоз встречается довольно часто даже у здоровых людей при интенсивной мышечной работе. У нетренированных людей лактоацидоз при физической работе возникает как следствие относительного недостатка кислорода в мышцах и развивается достаточно быстро. Компенсация осуществляется путём гипервентиляции.
При некомпенсированном лактоацидозе содержание лактата в крови увеличивается до 5 ммоль/л (в норме до 2 ммоль/л). При этом pH крови может составлять 7,25 и менее (в норме 7,36-7,44).
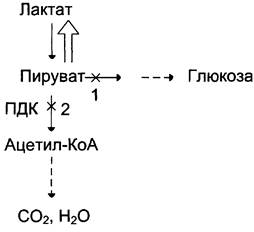
Повышение содержания лактата в крови может быть следствием нарушения метаболизма пирувата (рис. 7-51).
Рис. 7-51. Нарушения метаболизма пирувата при лактоацидозе. 1 — нарушение использования пирувата в глюконеогенезе; 2 — нарушение окисления пирувата.
Так, при гипоксии, возникающей вследствие нарушения снабжения тканей кислородом или кровью, уменьшается активность пируватдегидрогеназного комплекса и снижается окислительное декарбоксилирование пирувата. В этих условиях равновесие реакции пируват лактат сдвинуто в сторону образования лактата. Кроме того, при гипоксии уменьшается синтез АТФ, что, следовательно, ведёт к снижению скорости глюконеогенеза — другого пути утилизации лактата. Повышение концентрации лактата и снижение внутриклеточного pH отрицательно влияют на активность всех ферментов, в том числе и пируваткарбоксилазы, катализирующей начальную реакцию глюконеогенеза.
Возникновению лактоацидоза также способствуют нарушения глюконеогенеза при печёночной недостаточности различного происхождения. Кроме того, лактоацидозом может сопровождаться гиповитаминоз В1, так как производное этого витамина (тиаминдифосфат) выполняет коферментную функцию в составе ПДК при окислительном декарбоксилировании пирувата (см. раздел 6). Дефицит тиамина может возникать, например, у алкоголиков с нарушенным режимом питания.
Итак, причинами накопления молочной кислоты и развития лактоацидоза могут быть:
• активация анаэробного гликолиза вследствие тканевой гипоксии различного происхождения;
• поражения печени (токсические дистрофии, цирроз и др.);
• нарушение использования лактата вследствие наследственных дефектов ферментов глюконеогенеза, недостаточности глюкозо-6-фосфатазы;
• нарушение работы ПДК вследствие дефектов ферментов или гиповитаминозов;
• применение ряда лекарственных препаратов, например, бигуанидов (блокаторы глюконеогенеза, используемые при лечении сахарного диабета).
В. Синтез глюкозы из аминокислот
В условиях голодания часть белков мышечной ткани распадается до аминокислот, которые далее включаются в процесс катаболизма. Аминокислоты, которые при катаболизме превращаются в пируват или метаболиты нитратного цикла, могут рассматриваться как потенциальные предшественники глюкозы и гликогена и носят название гликогенных. Например, оксалоацетат, образующийся из аспарагиновой кислоты, является промежуточным продуктом как нитратного цикла, так и глюконеогенеза.
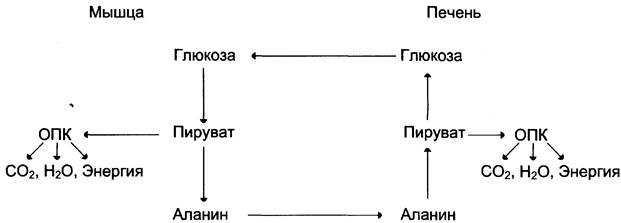
Из всех аминокислот, поступающих в печень, примерно 30% приходится на долю аланина. Это объясняется тем, что при расщеплении мышечных белков образуются аминокислоты, многие из которых превращаются сразу в пируват или сначала в оксалоацетат, а затем в пируват. Последний превращается в аланин, приобретая аминогруппу от других аминокислот. Аланин из мышц переносится кровью в печень, где снова преобразуется в пируват, который частично окисляется и частично включается в глюконеогенез. Следовательно, существует следующая последовательность событий (глюкозо-аланиновый цикл): глюкоза в мышцах —> пируват в мышцах —> аланин в мышцах —> аланин в печени —> глюкоза в печени —> глюкоза в мышцах (рис. 7-52). Весь цикл не приводит к увеличению количества глюкозы в мышцах, но он решает проблемы транспорта аминного азота из мышц в печень и предотвращает лактоацидоз.
Рис. 7-52. Глюкозо-аланиновый цикл.
Г. Синтез глюкозы из глицерола
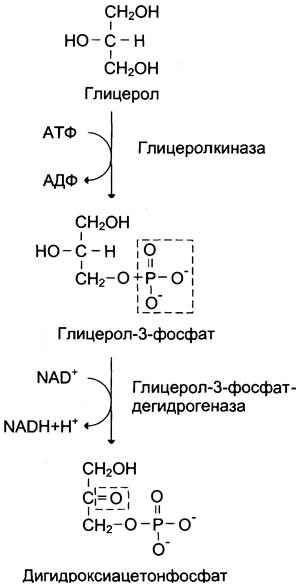
Глицерол образуется при гидролизе триацил- глицеролов, главным образом в жировой ткани. Использовать его могут только те ткани, в которых имеется фермент глицерол киназа, например, печень, почки. Этот АТФ-зависимый фермент катализирует превращение глицерола в α-глицерофосфат (глицерол-3-фосфат). При включении глицерол-3-фосфата в глюконеогенез происходит его дегидрирование NAD-зaвисимой дегидрогеназой с образованием дигидроксиацетонфосфата (рис. 7-53), который далее превращается в глюкозу.
Рис. 7-53. Превращение глицерола в дигидроксиацетон- фосфат.
Биологическая библиотека - материалы для студентов, учителей, учеников и их родителей.
Наш сайт не претендует на авторство размещенных материалов. Мы только конвертируем в удобный формат материалы, которые находятся в открытом доступе и присланные нашими посетителями.
Если вы являетесь обладателем авторского права на любой размещенный у нас материал и намерены удалить его или получить ссылки на место коммерческого размещения материалов, обратитесь для согласования к администратору сайта.
Разрешается копировать материалы с обязательной гипертекстовой ссылкой на сайт, будьте благодарными мы затратили много усилий чтобы привести информацию в удобный вид.
[youtube.player]Читайте также:
