
Аскарида первичноротая или вторичноротая
При описании развития живых организмов часто можно встретить термины "первичноротые" и "вторичноротые животные". Это все существующие многоклеточные, кроме кишечнополостных, которые различаются на эмбриональном уровне.
Развитие хордовых животных
Хордовые представлены тремя группами организмов:
- оболочниками, или личиночнохордовыми (Tunicata, Urochordata), которые имеют мешкообразное тело, окутанное оболочкой;
- бесчерепными, или головохордовыми (Acrania), живущими в морской глубине рыбообразными малоподвижными существами;
- позвоночными - представителями рыб, амфибий, змей, птиц, животных и человека.

История зарождения хордовых окутана таинственностью и вызывает множество предположений. И чтобы их конкретизировать, нужно разобраться, кто такие вторичноротые и первичноротые животные. Достоверно известно, что более 500 миллионов лет назад уже существовали головохордовые. Предшествующие этапы остаются загадками, поэтому возникают различные гипотезы происхождения живых существ.
Сущность первичноротых и вторичноротых животных
Многоклеточные представляют царство Животные. Первичноротые и вторичноротые - их прямые предки. Их словесные обозначения Protostomia и Deuterostomia образованы от особенностей формирования рта у эмбрионов. У первых он формируется из отверстия первичной кишки (бластопора), которая формируется у эмбриона. У вторых - в бластопоре развивается выводящее отверстие (анус), а ротовое отверстие прорезается на новом месте.
Классификация двусторонне симметричных организмов
Вторичноротые и первичноротые животные имеют свои классификации. К первичноротым относят:
- Моллюсков.
- Кольчатых червей.
- Сипункулидов - червеобразных морских животных, у которых выявлено наличие зачаточной системы кровообращения, выделительной системы и петлеобразной кишки с анусом со стороны спины.
- Эхиуридов - организмы, живущие в морских водах. У них тело цилиндрической формы с хоботком. Ротовое отверстие располагается с брюшинной стороны. От других представителей их отличает наличие слаборазвитой кровеносной системы и трубчатого кишечника, который заканчивается анусом.
- Пятиустков или язычковых, которые представляют собой червеобразных паразитических беспозвоночных с языковидным тельцем, похожих на членистоногих.
- Онихофоров или первичнотрахейных - представлены наземными хищниками, на длинном теле которых насчитывается до 43 пар ног.
- Тихоходок - мелких организмов с четырьмя парами конечностей.
- Членистоногих.

Вторичноротые животные, или дейтеростомы, представлены следующими типами животных:
- щетинкочелюстными;
- полухордовыми (надкишечнодышашие и крыложаберные);
- иглокожими;
- погонофорами;
- хордовыми;
- граптолитами (ископаемыми).

Этапы развития дейторостом непонятны. Предполагают, что их эволюция началась с многоклеточных беспозвоночных животных, которые никак не пересекается с первичноротыми. Другие считают, что предками вторичноротых являются низшие черви, которых причисляют к примитивным типам первичноротых.
В чем отличие первичноротых от вторичноротых
Кроме образования входного отверстия, представленного ртом, вторичноротые и первичноротые животные различаются рядом иных характеристик:
- По ряду делений яиц: у первых они радиальные, направленные по лучевой, а у вторых - спиральные (неравномерные).
- Способами закладывания целомы (полости): у первичноротых развитие стенок вторичных полостей начинается с клеточного деления, а у вторичноротых формируются из выступов кармашков кишечника эмбриона.
- Последующим генезисом головного мозга: у первичноротых - он развивается во взрослое состояние, а у вторых - уменьшается и закладывается в новой области.
Поэтому вторичноротых еще называют вторичномозговыми существами.

Ученые считают, что вторичноротые и первичноротые животные имеют одного общего прародителя, обитавшего в Эдикарском море более 500 миллионов лет назад. Средой обитания существа было морское дно, по которому он передвигался, работая ресничками, расположенными полосой на брюшке, и питался посредством щупальцев, которыми подбирал пищу. Возможно, позже задняя часть тела обособилась и прародители вторичноротых пользовались ею, чтобы на время закапываться в грунтовый слой.
Вторичноротые и первичноротые животные схожи по развитию и строению. Но также у них присутствует ряд отличий, благодаря чему их последователи отличаются между собой.
Первичноротые животные – это группа животных, у которых ротовое отверстие формируется на месте бластопора (первичного рта).
Основные характеристики первичноротых и вторичноротых животных
Группа первичноротых включает в себя членистоногих, моллюсков, а также червей. У представителей данной группы меоздерма формируется из телобластов. При этом полость тела образуется путем деления плотного тяжа клеток. Такой способ носит название схизоцельного.
Другая группа двустороннесимметричных животных называется вторичноротые (Deuterostomia). В эту группу входят хордовые и иглокожие. Дефинитивный рот у вторичноротых формируется не на месте бластопора, энетероцельным способом, и происходит становление целома (вторичной полости тела).
Также первично- и вторичноротые различаются по типам дробления. Что касается вторичноротых, то бластомеры у них располагаются по квартетам один над другим. При этом анимально-вегетативная ось располагается радиально, поэтому тип симметрии является радиальным. Что касается первичноротых, то у них наблюдается спиральный тип дробления.
Попробуй обратиться за помощью к преподавателям
Спиральный тип дробления – это дробление, при котором бластомеры вышележащего квартета располагаются между бластомерами нижележащего. Кроме того, бластомеры первичноротых обладают меньшей способностью к регуляции развития, чем бластомеры вторичноротых.
Бластопор – это отверстие, с помощью которого первичная кишка (гастроцель) сообщается с окружающей средой на такой стадии дробления, как гаструла.
В процессе развития у первичноротых животных бластопор обычно превращается в рот или образует рот и анальное отверстие, а у вторичноротых образует анальное отверстие.
Помимо механизмов образования и месторасположения рта первичноротые и вторичноротые животные отличаются целым рядом эмбриологических признаков:
- характер дробления (особенности описаны выше);
- способ закладки целома. Первичноротые продуцируют стенки вторичной полости тела из двух клеток, а вторичноротые за счет карманов эмбрионального кишечника при выпячивании;
- судьба зачатка первичного мозга: для первичноротых характерно развитие мозга взрослых форм из первичного мозга, а у вторичноротых он практически всегда редуцируется и обновленный нервный центр возникает в новом месте. Таким образом, вторичноротых животных можно называть и вторично мозговыми.
Задай вопрос специалистам и получи
ответ уже через 15 минут!
Как уже отмечалось ранее первичноротые и вторчиноротые животные имеют собственные классификации. Для полного понимания особенностей строения вторчиноротых и первичноротых животных необходимо привести пример более детальной классификации указанных групп. Среди первичноротых выделяют:
- моллюсков;
- кольчатых червей;
- сипункулидов – это червеобразные морские животные с зачаточной системой кровообращения, выделительной системы, кишечника в виде петли с анальным отверстием со стороны спины;
- эхиуридов – морских животных с телом цилиндрической формы и хоботком, у которой ротовое отверстие располагается со стороны брюшины. Кишечник имеет трубчатую форму и заканчивается анусом;
- пятиусток – язычковх животных, которые имею червеобразную форму тела и ведут паразитический образ жизни:
- первичнотрахейных хищников с длинным телом, у которых до 43 пар ног;
- тихоходок – небольших животных с четырём парами конечностей.
Кроме того, к первичноротым животным относят членистоногих.
Что касается вторичноротых животных, то среди них отмечают:
- щетинкочелюстных;
- полухордовых (крыложаберных и надкишечнодышащих);
- иглокожих;
- хордовых;
- ископаемых граптолитов.
Особенности первичноротых, а также вторичноротых животных в контексте сравнения представлены в таблице.
Рисунок 1. Особенности первичноротых и вторичноротых животных. Автор24 — интернет-биржа студенческих работ
Многие ученые придерживаются мнения о том, что вторичноротые и первичноротые животные происходят от одного предка. Считается, что он обитал в Эдикарском море примерно 500- 600 млн лет назад. Это существо вело морской образ жизни и передвигалось по субстрату с помощью ресничек. Они располагались на брюшной стороне тела в одну линию. Питание осуществлялось с помощью щупалец, которым пища захватывалась достаточно активно. Гипотеза также содержит информацию о том, что задняя часть тела животного обособилась и служила для данных предковых форм способом для закапывания в грунт.
Особенности хордовых животных по отношению к другим вторичноротым
Наконец, нельзя не отметить тот факт, что хордовых относят ко вторичноротым. Об этом говорят следующие факты: рот и анус хордовых животных формируются без связи с бластопором. Хордовые имеют типичное радиальное дробление яйца, вторичная полость тела (целом) формируется у них за счет выпячивания карманов первичного кишечника, а в нервной системе нет следов первичного мозга. Все эти признаки указывают на принадлежность хордовых к вторичноротым животным.
При этом они имеют целый ряд существенных отличий от других представителей данной группы. К ним относят следующие факты:
- бластопор хордовых находится на спинной стороне;
- нервная пластинка хордовых превращается не в брюшной нервный тяж, а в спинную трубку (за счет эмбриональной инверсии сторон тела);
- первая пара целомов диссметерична (правый передний целом меньше левого);
- отверстия целомов открываются на брюшной стороне;
- кровь по спинной стороне течет назад, а по брюшной вперед.
Все выявленные несоответствия означают тот факт, что предки хордовых претерпели изменения в теле, а именно подверглись инверсии. Морфологически брюшная сторона предков хордовых стала функционировать как физиологически спинная и наоборот.
Таким образом, подводя итог всему вышесказанному, можно сделать вывод о том, что вторичноротые и первичноротые животные очень сходны по своему строению и развитию. Но при этом у них присутствует ряд отличий, что делает их объектом пристального внимания научных исследований. Среди них необходимо, прежде всего, оценивать эволюционное разнообразие предковых форм первичноротых и вторчиноротых животных.
Так и не нашли ответ
на свой вопрос?
Просто напиши с чем тебе
нужна помощь
Вторичноротые (лат. Deuterostomia) — подраздел (иногда надтип) билатеральных животных. К ним относятся в том числе и наиболее прогрессивные животные — позвоночные (подтип хордовых).
Основные особенности вторичноротых:
- Дробление радиальное.
- Целомическая мезодерма закладывается энтероцельным способом.
- На месте бластопора прорывается анус (иногда бластопор непосредственно превращается в анус), рот не связан с бластопором (отсюда название - "вторичноротые").
- Ресничные личинки, как правило, имеются.
- Ресничные эпителии имеются.
- Имеется внутренний скелет – мезодермальный или энтодермальный.
- Вторичная полость тела подразделена на три отдела, иногда метамерная.
- Нервная система в виде эктодермальных нервных тяжей или нервных трубок, надглоточного ганглия нет.
Еще один характерный для вторичноротых признак — жаберные щели. Так называются парные отверстия, которыми передний отдел кишечника открывается наружу. Функция жаберных щелей — или дыхательная (в их стенках идет газообмен), или питательная (служат для отфильтровывания из воды мелких пищевых частиц), или обе сразу. Жаберные щели есть абсолютно у всех хордовых, хотя у наземных позвоночных — только на зародышевых стадиях. Есть жаберные щели и у полухордовых. У червеобразных представителей этого типа (кишечнодышащие) они развиты очень хорошо, у сидячих (крыложаберные) могут исчезать, но это наверняка вторичное состояние. Наконец, у современных иглокожих жаберных щелей нет [а чем они дышат?], но существуют палеонтологические данные, показывающие, что у их древних родичей таковые были. Таким образом, жаберные щели — это общая черта всех основных групп вторичноротых. Вероятно, они уже были у общего предка этой эволюционной ветви.
По-видимому, уже в самом начале эволюции вторичноротых в их теле сложилась система протока воды, входившей через рот и выходившей через жаберные щели, попутно отдавая пищевые частицы.
Можно предложить для объяснения происхождения жаберного аппарата следующий внутренне непротиворечивый эволюционный сценарий:
- Древние предки кишечнодышащих (и заодно всех вторичноротых) были сегментированными животными. В каждом туловищном сегменте имелась (а) пара целомодуктов и (б) пара печеночных карманов кишки. Целомодукты служили для выделения, карманы кишки — для пищеварения, между собой они никак не были связаны. Никаких жаберных щелей тогда еще не существовало.
- Потом в нескольких передних сегментах туловища произошло слияние целомодуктов с карманами кишки. Тем самым появилась возможность выводить продукты обмена веществ не только прямо наружу, но и через кишечник; это могло быть важно для роющего животного, обитавшего в норе, где выделять жидкость вбок не очень-то удобно. Но в результате карманы кишки получили выход наружу через дистальные (удаленные от середины тела) части целомодуктов. Это и можно считать моментом образования жаберных щелей, которые по определению являются каналами, связывающими кишку с внешней средой.
- В дальнейшем жаберные структуры приобрели дыхательную функцию, которая и стала у них главной. Связь с выделительной системой сохранил только самый передний жаберный мешок, в который впадает воротничковый целомодукт. Туловищные целомодукты исчезли. Возникла та ситуация, которую мы видим у современных кишечнодышащих.
- У хордовых выделительные органы окончательно потеряли прямую связь с жаберными щелями. У ланцетника выделительные трубочки впадают в атриальную полость, а у позвоночных — в совершенно самостоятельный выводной проток, который на зародышевой стадии называется вольфовым каналом.
Разделы страницы о вторичноротых животных:
- Эволюционное место и происхождение вторичноротых животных
- Вероятные ископаемые предки вторичноротых организмов
- Обзоры и статьи о вторичноротых многоклеточных
Также читайте о происхождении и эволюции хордовых животных и о самых примитивных хордовых - ланцетниках.
Эволюционное место и происхождение вторичноротых животных
 |
 |
Ксенотурбелла (лаь. Xenoturbella) — род морских червеобразных животных до 4 см в длину, выделяемый в отдельный тип Xenoturbellida. найдены у побережья Швеции (на глубине 60—100 м во фьордах), Шотландии и Исландии. Известно 6 видов. Первые экземпляры Xenoturbella были обнаружены в шведских водах Балтики в 1915 году биологом Сикстеном Боком (1884—1946), но описаны только в 1949 году Эйнаром Вестбладом под названием Xenoturbella bocki. Последний отнес их к Acoelomorpha, которые были тогда в составе плоских червей. Известны также по имени "лиловый носок", потому что похожи именно на брошенный на пол носок.
Существа эти настолько необычны, что в течение 60 лет ученые не могли определить, к какому именно виду или роду они принадлежат. Классификация и сейчас затруднительна, но предварительно их считают самыми примитивными вторичноротыми. Из-за того, что они питаются моллюсками, а в личиночной стадии паразитируют на них, их тоже сначала считали моллюсками.
Сквозной пищеварительный тракт отсутствует - имеется только мешкообразная кишка без заднего прохода, образующая единственную полость тела. Чепм кушает, тем и выделяет (как гидра). Причем, ротовое отверстие довольно маленькое. Ни зубов ни сосательного хоботка нет. У них также нет централизованного мозга или нервного узла. Нервная система, расположена под эпидермисом, имеет диффузный характер. Имеется орган равновесия - статоцист. Половой системы тоже нет, но обнаружены гаметы - репродуктивные клетки, имеющие гаплоидный набор хромосом.
 |
Исследуя породы необычных сланцев в провинции Шаньси, которые сформировались во время "кембрийского взрыва" (около 520-510 миллионов лет назад), китайские и американские палеонтологи нашли останки необычного животного — небольшой овальной структуры длиной в один миллиметр, похожей на мешок с "зубастыми" краями. Оно было названо Saccorhytus coronarius, что переводится с латыни как "сморщенный мешок с короной".
Эти организмы, как предполагают ученые, обладали неким подобием жабр, следы которых можно заметить в виде конических структур на его коже. Через них "мешок" выпускал воду, которую проглатывал вместе с пищей. Впоследствии эти конусы могли превратиться в жаберные дуги, а затем — в челюсти и нос первых рыб.
Все билатерально-симметричные организмы делятся на две группы - первичноротые и вторичноротые, и раньше, чем приступить к анализу проблемы происхождения хордовых, следует выяснить, к какой из этих двух групп принадлежат хордовые. Названия "первичноротые" (Protostomia) и "вторичноротые" (Deuterostomia) произошли от способа развития ротового отверстия в эмбриогенезе. Так, у первичноротых животных бластопор (возникающее в эмбриональном развитии отверстие первичного кишечника) частично или полностью переходит в ротовое отверстие. У вторичноротых (например, у иглокожих) бластопор становится анальным отверстием, а рот прорывается заново на переднем конце личинки. В других случаях бластопор закрывается, а рот и анус открываются заново.
Помимо судьбы рта, первичноротые и вторичноротые отличаются другими эмбриологическими особенностями. Во-первых, это характер дробления оплодотворенного яйца: у первичноротых оно спиральное, а у вторичноротых - радиальное (рис. 1.2). Другое отличие состоит в способе закладке целома (вторичной полости тела): у первичноротых стенки вторичной полости тела происходят от двух клеток, а у вторичноротых за счет выпячивания карманов эмбрионального кишечника (рис. 1.2). Третьим важнейшим различием между первично- и вторичноротыми является судьба зачатка первичного мозга: у первичноротых он дает начало мозгу взрослых форм, а у вторичноротых всегда редуцируется, так что новый нервный центр возникает в другом месте заново. Вторичноротые - это вторичномозговые животные.
Рот и анус хордовых животных формируются без связи с бластопором. Хордовые имеют типичное радиальное дробление яйца, вторичная полость тела (целом) формируется у них за счет выпячивания карманов первичного кишечника, а в нервной системе нет следов первичного мозга. Все эти признаки указывают на принадлежность хордовых к вторичноротым животным. Помимо хордовых к ним относятся погонофоры, полухордовые и иглокожие (рис. 1.2.). В состав первичноротых входят плоские и круглые черви, кольчатые черви, моллюски, членистоногие (рис. 1.2).

Рис. 1.2 Особенности эмбрионального развития первичноротых и вторичноротых животных.
Сравнение хордовых с другими вторичноротыми
Хотя хордовые - это вторичноротые животные, все же их развитие имеет ряд особенностей. На рис. 1.4 представлены соответствующие стадии развития одной из групп вторичноротых - погонофор и низших хордовых - асцидий.
Анимальный полюс яйца погонофор, на котором располагается ядро яйцеклетки, соответствует спинной стороне эмбриона, а вегетативный, на котором концентрируется желток, - брюшной. У низших хордовый ситуация обратная: анимальный полюс соответствует брюшной стороне, а вегетативный - спинной. Бластопор (то есть та область эмбриона, где впячивается энтодерма и формируется первичный кишечник) у погонофор соответствует брюшной стороне личинки, а у низших хордовых - спинной. Зачаток нервной системы, закладывающийся по линии замыкания бластопора, у погонофор становится брюшным нервным тяжем, а у хордовых - спинной нервной трубкой (рис. 1.4).

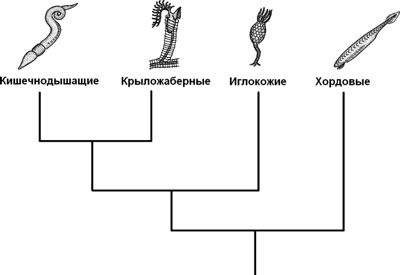
Рис. 1.3. Представители основных групп первичноротых и вторичноротых животных.
План строения низших хордовых также существенно отличается от планов строения других вторичноротых. Так, например, у всех вторичноротых передние целомы диссимметричны: левый передний целом больше правого, который обычно выполняет функцию околосердечной сумки. У хордовых ситуация обратная: правый передний целом больше левого. Целомы вторичноротых сообщаются с внешней средой отверстиями, открывающимися на спинной стороне и выполняющими осморегуляторные и половые функции. У хордовых эти отверстия располагаются на той стороне тела, которую мы традиционно считаем брюшной.
Кровеносная система у вторичноротых и хордовых устроена по общему плану и состоит из двух главных сосудов: спинного и брюшного. У вторичноротых, как и всех других беспозвоночных, кровь по спинному сосуду течет вперед, а по брюшному назад. У хордовых направление тока крови обратное: по спинному сосуду кровь течет назад, а по брюшному - вперед. Сердце у вторичноротых лежит на спинной стороне, а у хордовых - на брюшной.
Суммируем все эти отличия.
1. Бластопор хордовых соответствует не брюшной, а спинной стороне. 2. Нервная пластинка хордовых, закладывающаяся по месту замыкания бластопора, превращается не в брюшной нервный тяж, а в спинную нервную трубку. 3. Диссимметрия первой пары целомов имеет обратный знак в сравнении с другими вторичноротыми: правый передний целом у хордовых больше левого. 4. Отверстия целомов у хордовых открываются не на спинной, а на брюшной стороне. 5. Кровь у хордовых по брюшной стороне течет вперед, а по спинной - назад, и сердце располагается не на спинной, а на брюшной стороне. Что же означают эти несоответствия? Только одно: предки хордовых в своем эволюционном развитии испытали инверсию сторон тела, то есть перевернулись и стали передвигаться на морфологически спинной стороне, которая стала функционировать как физиологически брюшная. Морфологически брюшная сторона предков хордовых стала функционировать как физиологически спинная.

Рис. 1.4. Особенности эмбрионального развития погонофор и низших хордовых
Если мы примем этот вывод, все перечисленные выше несоответствия исчезнут: бластопор хордовых станет соответствовать их исходно брюшной стороне, нервная трубка будет располагаться на морфологически брюшной стороне, диссимметрия передних целомов приобретет нормальный для вторичноротых характер, отверстия целомов окажутся на морфологически спинной стороне, и направление тока крови, так же как положение сердца, окажется типичным для вторичноротых. Переворот на спинную сторону мог стать тем важнейшим эволюционным событием, которое определило формирование плана строения хордовых животных и его коренное отличие от планов строения вторичноротых и других беспозвоночных.
Если внимательно рассмотреть ранний эмбриогенез целомических многоклеточных, то мы обнаружим несколько фундаментальных черт, которые предполагают разделение их на две большие группы форм животных: ветвь кольчатых червей - членистоногих - моллюсков ( КЧМ ) и ветвь иглокожих - хордовых ( ИХ ) ( рис. 4.3 ). У кольчатых червей - членистоногих - моллюсков первые деления зигот (деления дробления) имеют спиральный характер ( рис. 4.1А(а) ). Слои дочерних клеток (бластомеров) выстраиваются со сдвигом относительно друг друга. Проспективная судьба клеток таких зародышей детерминирована. Если одна из них удалена, соответствующая часть организма не сформируется. Напротив, в линии развития иглокожих - хордовых зигота претерпевает радиальное дробление ( рис. 4.1А(б) ), а слои клеток расположена строго в один над (под) другим. Судьбы бластомеров на этой стадии строго не детерминированы. Если один из них удален, остальные способны это компенсировать, так что формируется нормальный зародыш.
Далее, две эти большие ветви демонстрируют глубокие различия путей формирования целома (полости тела). В линии КЧМ это происходит в результате расщепления мезодермы ( шизоцель ). В линии ИХ это происходит в результате отшнуровки выпячиваний кишки ( энтероцель ). Наконец, важное различие - в том, как формируется рот. Сфера из клеток, образующаяся в результате делений дробления оплодотворенной яйцеклетки - бластула - затем инвагинирует, формируя первичную кишку - архэнтерон. Этот процесс называется гаструляцией. В ветви КЧМ отверстие первичной кишки - бластопор - в конце концов развивается в рот взрослой особи. Животные, у которых рот развивается таким образом, относятся в первичноротым . Напротив, в ветви ИХ рот развивается иначе. В этом случае новое отверстие в конце концов возникает на конце, противоположном бластопору, а бластопор преобразуется в анус ( рис. 4.1А(б) ). Животные с таким типом развития относятся к вторичноротым(поскольку рот развивается у них "вторично"). Эти два термина - первичноротые и вторичноротые - обычно употребляются для обозначения двух упомянутых выше крупных ветвей развития.
Одно из двух основных подразделений животного царства ( Metazoa ), у представителей которого ротовое отверстие взрослого животного возникает на месте зародышевого рта.
Особенности раннего развития - одна из важнейших групп признаков, которые позволяют установить родственные отношения животных. Эти признаки часто мало изменяются в ходе эволюции. Недаром бластула игаструла - стадии, имеющие много общего у самых разных групп животных.
К первичноротым относятся плоские, круглые и кольчатые черви , моллюски и членистоногие . Для них характерно детерминированное дробление и спиральное расположение бластомеров ( рис. 154 ). Рот у них обычно образуется на месте бластопора (отсюда и название группы), а анальное отверстие, если оно есть, образуется в другой части гаструлы ( рис. 154 , III). Основная часть мезодермы у первичноротых образуется из двух крупных клеток, лежащих в задней части тела по бокам кишечника ( рис. 154 , II).
1)Важное различие первичноротых и вторичноротых в том, как формируется рот. Сфера из клеток, образующаяся в результате делений дробления оплодотворенной яйцеклетки - бластула - затем инвагинирует, формируяпервичную кишку -архэнтерон . Этот процесс называется гаструляцией . В ветви КЧМ (кольчатых червей -членистоногих - моллюсков) отверстие первичной кишки - бластопор - в конце концов развивается в рот взрослой особи. Животные, у которых рот развивается таким образом, относятся в первичноротым. У Protostomia первичный рот (бластопор) зародыша (именно гаструлы) переходит в рот взрослого животного или же дефинитивный рот образуется на месте первичного рта ( рис. 4.1А(б) ). Мезодерма формируется, как правило, телобластическим способом, т.е. из пары специальных клеток зародыша. К первичноротым относятся типы кольчатых червей (Annelida), моллюсков (Mollusca), членистоногих (Arthropoda) и онихофор (Onychophora).
Накапливаются молекулярно-биологические данные, дающие основания полагать, что первичноротые очень рано разделились на две крупных группы - Ecdyozoa (с жестким покровом) и Lophotrochozoa (без жесткого покрова).
2)Одно из двух основных подразделений животного царства ( Metazoa ), у представителей которого в онтогенезе происходит замыкание рта зародыша и возникновение ротового отверстия взрослого животного на ином месте.
Важное различие первичноротых и вторичноротых в том, как формируется рот. Сфера из клеток, образующаяся в результате делений дробления оплодотворенной яйцеклетки - бластула - затем инвагинирует, формируя первичную кишку - архэнтерон. Этот процесс называется гаструляцией. У животных ветви ИХ (иглокожих -хордовых) рот развивается так: новое отверстие в конце концов возникает на конце, противоположном бластопору, а бластопор преобразуется в анус ( рис. 4.1А(б) ). Животные с таким типом развития относятся к вторичноротым (поскольку рот развивается у них "вторично").
\типы вторичноротых! К вторичноротым относятся пять типов животных:
тип иглокожие, Echinodermata: морские звёзды, морские ежи и др.,
§ тип щетинкочелюстные, Chaetognatha,
§ тип ксенотурбеллиды, Xenoturbellida,
§ тип полухордовые, Hemichordata,
§ тип хордовые, Chordata: в том числе позвоночные (млекопитающие, рыбы, птицы и др.).
§ Ранее существовали сомнения насчёт щетинкочелюстных, но в последнее время их принято также относить к вторичноротым [источник не указан 271 день] .
§ Ещё один тип животных, возможно, относящийся к вторичноротым, пока имеет неясный статус — Ветуликолии (Vetulicolia).
Дата добавления: 2015-04-19 ; просмотров: 2600 . Нарушение авторских прав
Читайте также:
- Энтеробиоз по методу рабиновича в инвитро
- Гонорея сифилис хламидиоз при оральном сексе
- От каких причин может показать в крови наличие сифилиса
- Какой сифилид является признаком злокачественного течения сифилиса
- Какая смертность при вич инфекции
 Пожалуйста, не занимайтесь самолечением!При симпотмах заболевания - обратитесь к врачу.
Пожалуйста, не занимайтесь самолечением!При симпотмах заболевания - обратитесь к врачу.