
У кишечной палочки атф синтезируется
Автор текста – Анисимова Елена Сергеевна.
Авторские права защищены. Продавать текст нельзя.
Курсив не зубрить.
Параграф 23:
Синтез АТФ - субстратное и окислительное фосфорилирование.
см. также 22, 20 и 21, 25.
Сокращения:
СФ – субстратное фосфорилирование,
ОФ – окислительное фосфорилирование,
ММП – межмембранное пространство митохондрий.
Содержание параграфа.
23. 1. Для чего нужен синтез АТФ клеткам.
23. 2. Фосфорилирование АДФ – синтез АТФ.
23.3. Чем отличаются СФ и ОФ.
23.4. Источники энергии для ОФ – прямой и непрямой.
23.5. Субстратное фосфорилирование.
23.5.1. Механизм СФ на примере с сукциниломКоА их ЦТК.
23.5.2. Механизм СФ на 2-х примерах реакций гликолиза.
23.6. М е х а н и з м О Ф .
23.6.1. О транспорте протонов из матрикса
23.6.2. О транспорте протонов в матрикс
23.7. На что тратится энергия, кроме синтеза АТФ.
23.8. Градиент протонов – форма конвертируемой энергии.
23.8.1. Как создать ЭХП протонов.
23.8.2. Два компонента ЭХП протонов.
23.9. Сопряжение и разобщение
окисления и фосфорилирования
(окислительного фосфорилирования).
23.10. Разобщение ОФ.
23.11. С р а в н е н и е блокаторов и разобщителей.
23.12. Действие ЙТ на работу ДЦ и на разобщение (п.104) –
Главное.
СФ и ОФ – это способы синтеза АТФ.
23.1. Для чего нужен синтез АТФ клеткам.
При использовании АТФ в качестве источника энергии
АТФ превращается в АДФ и фосфат (Н3РО4).
(Иногда просто в АДФ – см. киназные реакции;
даже если АТФ расщепляется на АМФ и дифосфат,
то далее АМФ превращается в АДФ
за счет получения фосфата от АТФ: АМФ + АТФ ; 2 АДФ).
Чтобы [АТФ] в клетке поддерживалась на необходимом уровне,
нужно постоянно превращать АДФ в АТФ.
Иначе все молекулы АТФ превратятся в АДФ,
а без АТФ клетка умирает (гипоэнергетический шок).
Примеры ситуаций, когда клетки и организм в целом
умирают от дефицита АТФ:
- смерть от отсутствия кислорода в воздухе,
- смерть от прекращения работы дыхательной системы,
- смерть из-за нарушения доставки кислорода в ткани при анемии,
- смерть при приеме цианистого калия и т.д..
Если бы в этих ситуациях клетки не теряли бы возможность синтезировать АТФ,
то эти ситуации не приводили бы к смерти.
23. 2. Фосфорилирование АДФ – синтез АТФ.
Для синтеза АТФ из АДФ
нужно к АДФ присоединить фосфат,
то есть фосфорилировать АДФ.
Как синтезируется АДФ, здесь не разбирается.
См. вопрос о синтезе пуриновых нуклеотидов в п.72.
Нужны Р-5-Ф из ПФП и некоторые аминокислоты в качестве сырья для синтеза АДФ.
23.3. Чем отличаются СФ и ОФ.
СФ и ОФ отличаются по источникам энергии,
которая тратится на образование макроэргической связи
между присоединяемым фосфатом и АДФ.
При СФ источником энергии для фосфорилирования АДФ
является расщепление макроэргической связи другого вещества,
а при ОФ источником энергии для синтеза АТФ
является перенос электронов в дыхательной цепи.
Хоть и не напрямую, как будет показано далее.
Процесс назвали ОФ, потому что
при переносе электронов происходит окисление тех веществ,
от которых отщеплен электрон.
23.4. Источники энергии для ОФ – прямой и непрямой.
Непосредственным источником энергии для ОФ
является не перенос электронов,
а пассивный транспорт протонов через канал АТФ-синтАзы,
при котором тратится энергия ЭХП протонов – далее об этом подробнее.
Из-за того, что источником энергии для ОФ является ДЦ,
которая требует кислорода в качестве субстрата,
ОФ протекает только в присутствии кислорода
(то есть только в аэробных условиях).
Это значит, что в анаэробных условиях клетка не может получать энергию способом ОФ.
Поскольку СФ дает намного меньше АТФ, чем ОФ
(от одной молекулы глюкозы:
2 АТФ при СФ вместо 32 АТФ при СФ вместе с ОФ),
то в анаэробных условиях возникает острый дефицит АТФ (и угроза гибели)
в тех клетках, которые тратят много АТФ:
- в головном мозге,
- в коре почек,
- в миокарде и в других аэробных тканях.
Только эритроциты могут существовать без ОФ,
хотя в них и много кислорода.
(в ЭЦ ОФ не происходит, т.к. нет митохондрий).
23.5. Субстратное фосфорилирование.
Вещество, расщепление макроэргической связи которого дает энергию для синтеза АТФ,
называется первичным макроэргом.
АТФ при СФ называется вторичным макроэргом.
Образование
первичного макроэрга происходит
за счет энергии, которая выделяется при окислении другого вещества,
которое называется субстратом СФ
(то есть не просто субстратом,
а именно субстратом для процесса СФ);
обычно первичный макроэрг образуется из субстрата СФ – см. далее примеры.
23.5.1. Механизм СФ на примере с сукциниломКоА из ЦТК.
В ЦТК кетоглутарат превращается в сукцинилКоА,
при этом происходит окислительное декарбоксилирование кетоглутарата.
При окислении кетоглутарата выделяется энергия,
за счет которой образуется макроэргическая связь
между сукцинилом и коферментом А (КоА).
При расщеплении МС сукцинилаКоА выделяется энергия,
за счет которой образуется
между ГДФ и фосфатом,
в результате чего образуется ГТФ.
ГДФ и ГТФ – это вещества, аналогичные АДФ и АТФ,
и синтез ГТФ происходит путем фосфорилирования ГДФ
(в данном случае – способом СФ).
Затем ГТФ вступает в реакцию с АДФ
и отдает ему фосфат,
в результате чего АДФ превращается в АТФ, а ГТФ – в ГДФ.
23.5.2. Механизм СФ на 2-х примерах реакций гликолиза.
См. реакции и формулы в п.32.
1-й пример СФ из гликолиза.
В одной из реакций гликолиза
ФГА превращается в 1,3-бис/фосфо/глицерат,
при этом происходит окисление ФГА
и присоединение фосфата макроэргической связью.
При окислении ФГА
(точнее, промежуточного метаболита, связанного с ферментом)
выделяется энергия,
за счет которой образуется
с присоединяемым по 1-му положению фосфатом.
1,3-бис/фосфо/глицерата
выделяется энергия,
за счет которой образуется
между АДФ и фосфатом
(переносимым от 1,3-бис/фосфо/глицерата),
в результате чего образуется АТФ (вторичный макроэрг).
1-й пример СФ из гликолиза.
В другой реакции гликолиза
2-ФГ превращается в фосфо/енол/пируват (ФЕП),
при этом происходит отщепление воды,
а связь с фосфатом становится макроэргической.
фосфо/енол/пирувата
выделяется энергия,
за счет которой образуется
между АДФ и фосфатом (переносимым от ФЕПа),
в результате чего образуется АТФ.
Между СФ в ЦТК и СФ в гликолизе есть важное отличие:
СФ в ЦТК, как и весь ЦТК,
не происходит в отсутствие кислорода и вне митохондрий
(то есть в анаэробных условиях;
МХ нет в ЭЦ;
ЦТК не работает в анаэробных условиях,
потому что должен превращать НАД+ в НАДН для ДЦ,
но без кислорода ДЦ не работает,
не потребляет НАДН и не превращает НАДН в НАД+).
А СФ в гликолизе может происходить в отсутствие кислорода
и без митохондрий.
Поэтому СФ в гликолизе –
единственный источник АТФ для клеток,
когда в них нет кислорода :
при ишемии, гипоксии,
из-за жгута, спазма, низкого артериального давления и т.д..
Или если в клетках нет митохондрий, как в эритроцитах.
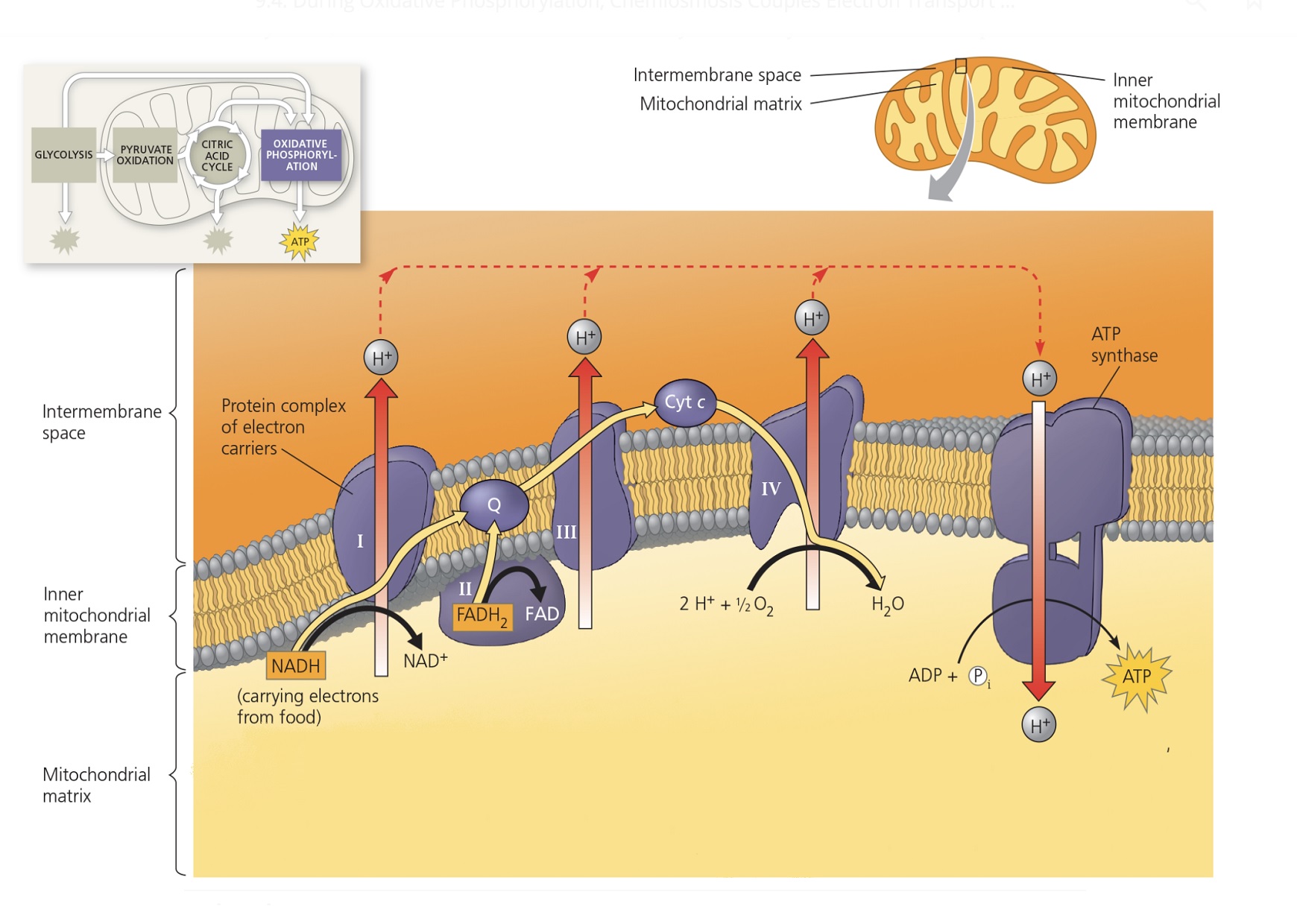
23.6. М е х а н и з м О Ф .
Уже говорилось, что при ОФ
АТФ синтезируется за счет энергии дыхательной цепи.
Поэтому говорят, что работа ДЦ сопряжена (связана) с синтезом АТФ
(с фосфорилированием АДФ).
Существует теория, которая описывает механизм ОФ
(то есть механизм сопряжения окисления в ДЦ и фосфорилирования АДФ).
Автор теории – Митчелл.
Названия теории ОФ – хемоосмотическая, протондвижущая.
Согласно теории Митчелла (ТМ),
энергия, выделяющаяся при переносе электронов по ДЦ,
не сразу тратится на синтез АТФ.
Сначала энергия ДЦ тратится на транспорт протонов (Н+)
из матрикса МХ в межмембранное пространство
через внутреннюю мембрану МХ
(против градиента протонов).
Затем протоны возвращаются в матрикс МХ
через канал фермента АТФ-синтазы,
и АТФ-синтаза синтезирует АТФ
за счет энергии, которая выделяется при транспорте протонов
(т.к. транспорт протонов в матрикс происходит по градиенту протонов,
пассивный, энергия не тратится, а выделяется).
Транспорт протонов через мембрану происходит с помощью белков-транспортеров. (Потому что липидный слой мембраны не проницаем для ионов.)
23.6.1. О транспорте протонов из матрикса
(в межмембранное пространство,
через внутреннюю мембрану МХ
с внутренней стороны внутренней мембраны
на внешнюю сторону внутренней мембраны;
Предполагается, что транспорт протонов из матрикса
осуществляется с участием белков ДЦ и кофермента Q.
При транспорте протонов из матрикса
происходит их накопление в ММП,
с внешней стороны мембраны оказывается больше протонов,
чем с внутренней стороны мембраны,
то есть возникает разница концентраций протонов
(градиент протонов)
с разных сторон мембраны.
Транспорт частиц против градиента
требует затрат энергии
(и поэтому называется активным).
Источником энергии для транспорта протонов из матрикса
является энергия, выделяющаяся при работе дыхательной цепи
(то есть при переносе электронов по ДЦ).
23.6.2. О транспорте протонов в матрикс.
Транспорт протонов в матрикс
происходит, в основном (о других вариантах позже)
через канал АТФ-синтазы.
При транспорте протонов в матрикс
[Н+] в матриксе увеличивается,
а в ММП [Н+] уменьшается,
в результате чего снижается разница концентраций протонов
с разных сторон мембраны.
Транспорт частиц по градиенту
НЕ требует затрат энергии
(и поэтому называется пассивным).
(Точнее, источником энергии является сам градиент, его снижение).
Более того, при транспорте частиц по градиенту
энергия выделяется
и может использоваться для совершения работы
(то есть процессов, требующих затрат энергии).
При транспорте протонов в матрикс энергия тоже выделяется.
АТФ-синтаза тратит энергию на синтез АТФ
(поэтому и называется АТФ-синтазой).
Как говорилось выше, способ синтеза АТФ при этом называется ОФ.
Работа, которая совершается за счет энергии,
выделяющейся при транспорте протонов в матрикс –
это синтез АТФ
(эта работа называется химической,
потому что заключается в обеспечении энергией химической реакции).
23.7. На что тратится энергия, кроме синтеза АТФ.
АТФ-синтаза использует на синтез АТФ
не 100% энергии, выделяющейся при транспорте протонов,
а около 40% энергии
(при разобщении ещё меньше – см. далее).
Остальная часть энергии (60%)
рассеивается в виде тепла
(это тепло называется первичным – см. далее)
и тратится на транспорты веществ через мембрану.
То есть на осмотическую работу.
23.8. Градиент протонов – форма конвертируемой энергии.
ЭХП протонов.
Поскольку пассивный транспорт протонов
может быть источником энергии для совершения работы,
то градиент протонов,
при наличии которого возможет пассивный транспорт протонов,
является формой конвертируемой энергии.
Эта форма энергии называется электро-химическим потенциалом (ЭХП) протонов
и обозначается ;µН+.
(Знак ; (дельта) означает, что есть разная концентрация;
µ означает, что это форма энергии).
Энергия, заключенная в ;µН+,
выделяется
и может быть использована при пассивном транспорте протонов,
то есть при транспорте протонов в область меньшей концентрации:
при этом разница концентраций протонов уменьшается
(градиент протонов уменьшается),
;µН+ уменьшается.
;µН+ возникает при появлении разных концентраций протонов
с разных сторонах внутренней мембраны митохондрий
(то есть при появлении градиента протонов).
И увеличивается при увеличении разницы [H+].
23.8.1. Как создать ЭХП протонов.
Для увеличения разницы [H+] (и создания ;µН+)
нужно транспортировать протоны
из области меньшей концентрации
в область большей концентрации
(то есть против градиента протонов),
то есть должен произойти активный транспорт протонов.
Для активного транспорта нужны затраты энергии.
Энергию для активного транспорта протонов
(и тем самым – для создания ;µН+) дает ДЦ.
23.8.2. Два компонента ЭХП протонов.
;µН+ называется ЭХП,
потому что в ;µН+ есть два компонента (и они отражены в названии): электрический и химический.
Электрический компонент ЭХП обусловлен тем, что
(при наличии на мембране градиента ионов)
с разных сторон мембраны
разные концентрации заряженных частиц
и в результате – разные заряды.
Другое название электрического компонента ЭХП – мембранный потенциал.
Химический компонент ЭХП
обусловлен тем, что
(при наличии на мембране градиента ионов)
с разных сторон мембраны
есть разные концентрации химического вещества.
Другое название химического компонента ЭХП –
концентрационный градиент.
Протоны создают кислую среду:
где больше протонов, там кислее среда и меньше рН.
Н+ имеет положительный заряд,
поэтому – чем больше протонов на данной стороне мембраны,
тем более положительно заряжена эта сторона мембраны.
Н+ больше в ММП (при работе ДЦ),
то есть на внешней стороне внутренней мембраны
(со стороны ММП).
На внешней стороне внутренней мембраны
больше положительно заряженных частиц.
Внешняя сторона внутренней мембраны МХ заряжена более положительно.
23.9. Сопряжение и разобщение
окисления и фосфорилирования
(окислительного фосфорилирования).
(Имеется в виду – окисления в ДЦ и фосфорилирования АДФ).
Когда энергия ДЦ используется для синтеза АТФ,
то говорят, что процесс ДЦ и процесс синтеза АТФ сопряжены
(окисление и фосфорилирование сопряжены).
При этом способ синтеза АТФ называется окислительным фосфорилированием.
Сопряжение имеет место тогда,
когда Н+, транспортированные из матрикса в ММП
за счет энергии ДЦ,
возвращаются из ММП обратно в матрикс через АТФ-синтазу.
23.10. Разобщение ОФ.
Это явление называется разобщением ДЦ и синтеза АТФ
или разобщением окислительного фосфорилирования.
При разобщении энергия возвращения протонов в матрикс
просто рассеивается в виде тепла,
что приводит к увеличению температуры тела.
Причины разобщения ОФ:
к разобщению приводят различные факторы,
которые позволяют протонам не проходить через АТФ-синтазу.
Пример – динитрофенол (применялся в промышлености)
и (возможно) иодтиронины.
Протоны могут не проходить через АТФ-синтазу,
если в мембране есть другой белок,
способный пропускать протоны
(протонный канал) –
примером такого белка является термогенин,
который есть в митохондриях бурой жировой ткани.
Разные факторы, которые приводят к разобщению
(являются причинами разобщения)
называются разобщителями.
Разобщителями являются жирные кислоты в буром жире
(способствуют работе термогенина),
некоторые микробные токсины
(дифтерийный).
Из всех названных разобщителей только жирные кислоты (в буром жире)
относят к физиологическим –
то есть разобщение ими не вредно для организма.
Остальные разобщители считаются патологическими.
(Об ЙТ далее).
23.11. С р а в н е н и е блокаторов и разобщителей.
Ещё раз про блокаторы ДЦ.
Нужно не путать действие блокаторов ДЦ и разобщителей ОФ.
Блокаторы прекращают работу ДЦ,
поэтому энергия ни запасается в виде АТФ (слабость),
ни рассеивается в виде тепла (озноб).
А разобщители не прекращают работу ДЦ
и даже (ЙТ) могут усиливать работу ДЦ.
23.12. Действие ЙТ на работу ДЦ и на разобщение (п.104) –
ЙТ активируют ДЦ
и при этом разобщают ОФ.
При небольшом избытке ЙТ (при начальном гипертиреозе)
соотношение эффектов активации ДЦ и разобщения ОФ таково,
что АТФ и тепла больше,
чем при нормальном количестве ЙТ
(это увеличивает энергичность и устойчивость к холоду).
Но при значительном избытке ЙТ (при запущенном гипертиреозе)
разобщение настолько сильное,
что количество АТФ становится меньше,
чем в норме
(при том, что ДЦ активнее, чем в норме).
При снижении количества ЙТ (при гипотиреозе) ДЦ менее активна,
энергии дает меньше,
при этом снижается количество и АТФ, и тепла.

Разработка предназначена для подготовки учащихся одиннадцатых классов к единому государственному экзамену по биологии. А так же может быть использована на уроках биологии учителями-предметниками для закрепления материала. Задания даны в виде тестов. Выполняя данные тесты, можно добиться успешной подготовки к сдаче ЕГЭ.
Верны ли следующие суждения
А. Делением размножаются только клетки, но не органоиды.
Б. Клетки размножаются делением. а вирусы нет.
1. Верно только А
2. Верно только Б
3. Верны обе формулировки
4. Обе формулировки не верны
А. При дигибидном скрещивании у гибридов каждая пара признаков наследуется независимо от других и дает с ними разные сочетания.
Б. Это закон чистоты гамет: в каждую гамету попадает только одна аллель из пары аллелей данного гена родительской особи.
Б. Пары альтернативных признаков не смешиваются и при образовании гамет по одному переходят в них в чистом виде.
А. Результатами эволюции можно считать разнообразие организмов и их приспособленность к условиям окружающей среды.
Б. Результатами эволюции считаются изоляция, борьба за существование и возникновение мутаций.
А. Наследственная изменчивость служит материалом для естественного отбора.
Б. Наследственная изменчивость способствует сохранению в популяции наиболее приспособленных особей
А. Многие белки выполняют каталитическую функцию.
Б. Некоторые гормоны имеют белковую природу.
Упрощение в строении животных, связанные с паразитическим образом жизни, относят к биологическому регрессу.
Б. Возникновение класса Насекомые, сопровождающееся повышением нового уровня их организации, - пример ароморфоза.
А. Генетический код триплетен – один триплет всегда кодирует только одну аминокислоту.
Б. Генетический код однозначен – в молекуле нуклеиновой кислоты одна аминокислота кодируется сочетанием трех последовательно расположенных нуклидов.
А. В результате митоза из одной диплоидной клетки образуется 4 гоплоидные.
Б. Редукция числа хромосом происходит в анафазе I.
А. Мутационная изменчивость носит случайный характер.
А. Митоз обеспечивает рост организма, регенерацию, лежит в основе бесполого размножения.
Б. Мейоз обеспечивает образование гамет, увеличивает генетическое разнообразие и лежит в основе комбинативной изменчивости.
Верны ли следующие суждения о фотосинтезе?
А. В световой фазе фотосинтеза происходит синтез Атор, образуются атомы водорода и молекулярный кислород.
Б. Световая фаза фотосинтеза происходит в строме хлоропласта.
Верны ли следующие суждения о химическом составе клетки?
А. Цинк относится к микроэлементам
Б. В состав хлорофилла входит железо.
Верны ли следующие суждения о строении и функциях органоидов?
А. Лизосомы относятся у двумембранным органоидам.
Б. У кишечной палочки АТФ синтезируются не в митохондриях.
Верны ли следующие суждения об обмене веществ?
А. Второй этап катаболизма – неполное окисление или бескислородный – протекает в цитоплазме..
Б. В результате гликолиза из одной молекулы глюкозы образуется 2 молекулы АТФ.
Верны ли следующие суждения об особенностях ферментов?
А. Ферменты – это специфические белки, которые присутствуют во всех живых организмах и играют роль биологических катализаторов.
Б. Ферменты в отличие от химических катализаторов способны катализировать несколько различных реакций.
Верны ли следующие суждения о приспособленности организмов?
А. К этиологическим приспособлениям относятся, н/п иглы ежа, дикобраза, колючки кактуса, акации, барбариса, которые защищают животных от врагов или препятствуют поеданию растений травоядными животными.
Б. Японские макаки зимой при наступлении холодов спускаются с гор к термальным источникам и подолгу греются в теплой воде – это пример физиологической адаптации.
Верны ли следующие суждения о происхождении жизни?
А. Невозможность самозарождения была доказана опытами Ф. Реди, Л. Пастера и др.
Б. Доказательствами инопланетного происхождения жизни служат НЛО, наскальные изображения летательных аппаратов
А. Молекулы ДНК, находящиеся в митохондриях и хлоропластах, не являются хранителями наследственной информации.
Б. Мутации, возникающие в молекулах ДНК, передаются на и-РНК в соответствии с правилами комплементарности.
А. Плазматическая мембрана состоит из лидидного бислоя и встроенных белков.
Б. Мембранные белки выполняют транспортные, рецепторные и другие функции.
А. В разных фазах мейоза клетка может нести диплоидный или гаплоидный набор хромосом.
Б. В течении всего митоза клетка кожи человека всегда диплоидна.
Верны ли следующие суждения:
№2: Верны обе формулировки
№3: Верно только А (борьба за существование и мутации – это факторы эволюции, а не результат)
№4 верно А (Б – неверно, т.к. сохранению в популяции наиболее приспособленных особей способствует естественный отбор, а наследственная изменчивость представляет собой материал для отбора).
№5 верны оба суждения
№6 Верно Б (А – неверно, т.к. упрощение в строении животных, связанное с паразитическим образом жизни, относят к дегенерации. А дегенерация, как и ароморфоз и идиоадаптация, тоже относится к биологическому процессу.
№7 Оба суждения неверны. А – генетический под триптолетен, т.е. в молекуле НК одна аминокислота кодируется сочетанием 3-х последовательно расположенных нуклеотидов.
Б – генетический код однозначен, т.е. один триплет (кодон) всегда кодирует только одну А.К.
Роль АТФ в энергетическом балансе
АТФ является основной молекулой энергии в живых системах. Он участвует в различных химических процессах, от химического биосинтеза до движения ресничек, сокращения мышц, активного транспорта молекул через клеточную мембрану или распространения электрического импульса через нервные волокна.
Производство и потребление энергии происходит через сеть ферментативных реакций (метаболизм). Центральным химическим соединением в метаболизме является аденозинтрифосфат (АТФ), который образуется в результате метаболических реакций (катаболизм) путем фосфорилирования аденозиндифосфата (АДФ) с образованием энергии около 30 кДж / моль (термодинамика). Большая часть АТФ производится в результате процессов в митохондриях (окислительного фосфорилирования). При использовании этой энергии в биологических процессах АТФ обычно гидролизуется до фосфата и АДФ (аденозинДИфосфата).
Вся биосинтетическая деятельность, как и многие другие клеточные действия, требует энергии. В основном для клеточных активностей источником энергии является именно АТФ. Молекула АТФ состоит из аденина, рибозы и трех фосфатных групп (ФГ). Последние с сильным отрицательным зарядом связаны двумя ковалентными высокоэнергетическими связями, которые при гидролизе выделяют относительно много энергии. Это демонстрирует важное свойство АТФ.
Поэтому без преувеличения можно сказать, что наиболее важным энергетическим соединением в клетке является трифосфат аденозина (АТФ), который по своему химическому составу является нуклеотидом.
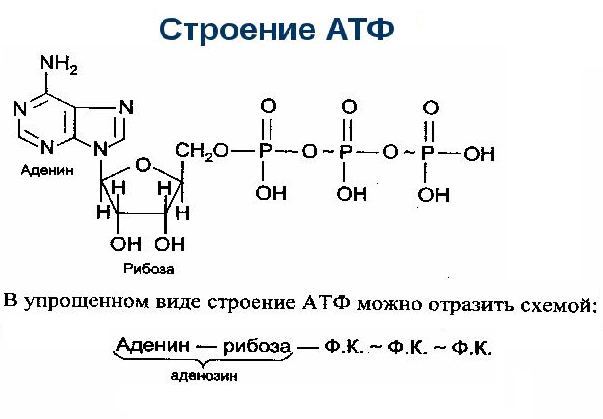
Молекула АТФ состоит из:
- - азотно-аденинового основания пурина;
- - пентозы, рибозы и моносахариды;
- - трех фосфатных групп, обозначенных как альфа, бета и гамма (начиная с рибозы).
Синтез АТФ в организме
АТФ чаще всего производится в митохондрии, в основном в результате расщепления глюкозы и жирных кислот в процессе, называемом окислительным фосфорилированием; разложение 1 молекулы глюкозы в митохондрии высвобождает 36 молекул АТФ. Также АТФ синтезируется в хлоропластах, при фотосинтезе в процессе фотосинтетического фосфорилирования.
АТФ не может храниться в качестве резерва, поэтому он расходуется после его синтеза путем дефосфорилирования с помощью фермента АТФазы. Две конечные фосфорные группы связаны богатыми энергией ковалентными связями. Когда эти связи разрушаются, высвобождается относительно большое количество энергии. Если от АТФ освободить один конец ФГ, то образуется аденозин дифосфат (АДФ), освободить другой - получится аденозинмонофосфат (АМФ).
Фосфорная группа, высвобождаемая из АТФ или АДФ, богата энергией и, связываясь с соединением, обогащает ее энергией (процесс, называемый фосфорилированием). Таким образом, энергия от АТФ используется в процессах анаболизма.
АТФ создается в качестве основного энергетического продукта процесса разложения пищевых ингредиентов в процессе окисления. Часть энергии, выделяемой в этих процессах, сохраняется в форме АТФ, а остальная часть используется в форме тепла. Полученный таким образом АТФ используется для взаимодействия со всеми типами клеток. Только около 1/3 АТФ расходуется на реакции анаболизма. Остальная энергия расходуется на движение, сокращение мышц, транспортировку вещества через клеточную мембрану и т. д.
Фосфорилирование, регенерация АТФ.
Восстановление (синтез) АТФ реализуется путем связывания ФГ сначала с АМФ, что приводит к АДФ, а затем из АТФ под контролем фермента АТФ-синтазы. Это возможно благодаря тепловым реакциям, в которых энергоемкие (анаболические) реакции связаны с энерговыделительными (катаболическими) реакциями. Энергия, выделяемая при катаболизме, используется для повторного синтеза АТФ из АДФ. Следовательно, система АТФ / АДФ служит универсальным способом обмена энергией, который балансирует между выделяемыми и потребляющими энергию реакциями.
Функциональные характеристики АТФ.
Химическая связь, представляющая собой сумму сил, которые удерживают вместе атомы в молекуле, является стабильной конфигурацией, и для разрыва старой связи и образования новой требуется энергия. Ферменты значительно снижают потребность в активации большого количества энергии, но для того, чтобы химические реакции происходили в живых организмах, необходимо, чтобы энергия связи в продуктах реакции всегда была меньше энергии связи реагентов.
Молекула, наиболее часто участвующая в тепловых реакциях, - АТФ. Внутренняя структура молекул АТФ отлично подходит для этой роли в живых системах. В лабораторных условиях при удалении третьей фосфатной группы образуются АДФ и фосфат, и выделяется около 7 ккал (30 кДж) на моль АТФ. Удаление второй фосфатной группы дает AMФ и фосфат, высвобождая такое же количество энергии.
Энергия, выделяемая при удалении фосфатных групп, не только возникает из высокоэнергетических связей, но также является результатом перераспределения орбит в молекулах АТФ или АДФ. Каждая фосфатная группа несет отрицательный заряд и поэтому имеет тенденцию отталкиваться от другой такой группы. Когда фосфатная группа удаляется, происходит изменение конфигурации электронов, в результате чего получается структура с меньшей энергией.
В живых системах АТФ также гидролизуется до АДФ. Гидролиз АТФ является, например, быстрым способом выработки тепла у животных, которые просыпаются от зимней спячки. Однако обычно конечный продукт не просто удаляется, а переносится через фермент (киназу) в другую молекулу (фосфорилирование). Эта реакция также передает часть энергии от высокоэнергетической связи фосфорилированному соединению, которое, таким образом, обогащается энергией при реакции.
Энергия, выделяемая в реакциях клеточного метаболизма, таких как расщепление глюкозы, используется для повторного синтеза АТФ из молекул АДФ. Основными механизмами синтеза АТФ в клетке являются окислительное фосфорилирование в процессе клеточного дыхания (на внутренней стороне митохондриальной мембраны) и фосфорилирование в процессе фотосинтеза.
Митохондрии
Митохондрии представляют собой мембранные органеллы, присутствующие в клетках практически всех эукариотических организмов. Митохондрии заключены в две мембраны: внешняя, находящаяся в контакте с цитоплазмой, и мембрана, ограничивающая внутреннюю часть митохондрий. Между этими двумя мембранами находится межмембранное пространство. Внутренняя часть митохондрий заполнена матриксом. Типичная эукариотическая клетка содержит около 2000 митохондрий.
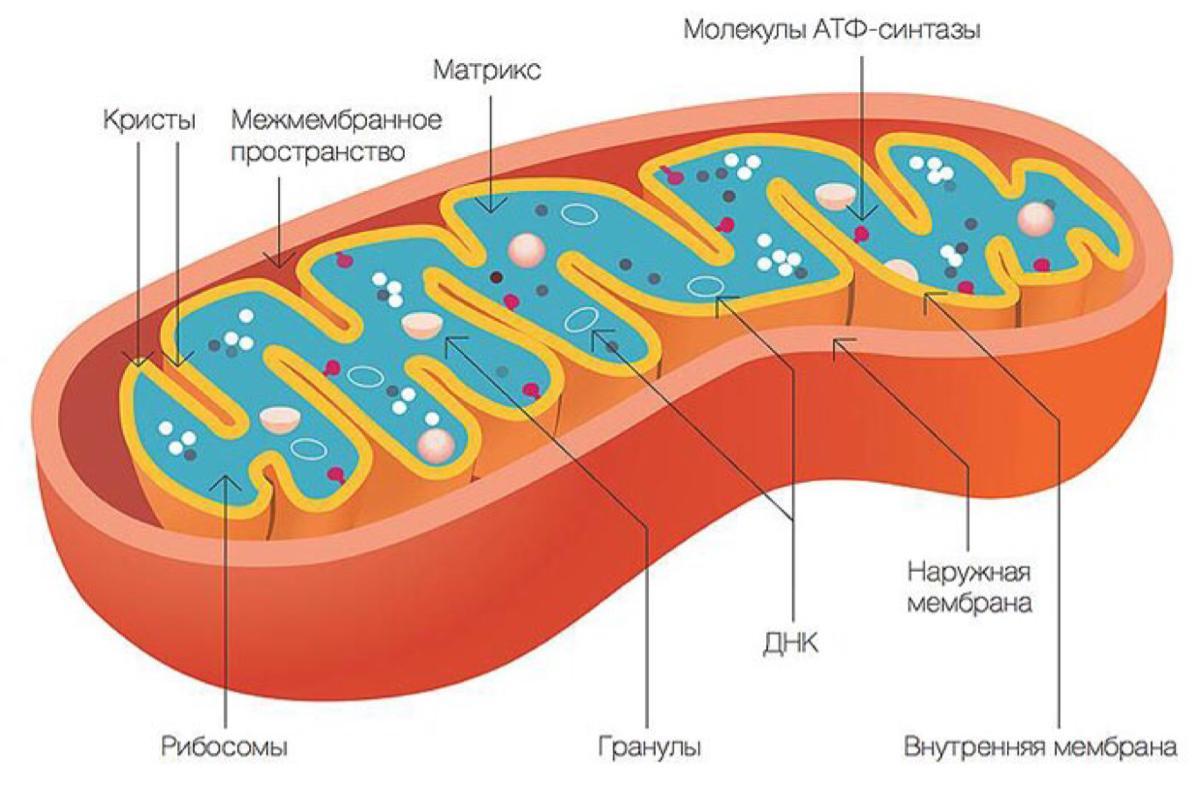
Внешняя митохондриальная мембрана определяет форму этой органеллы и, благодаря наличию каналообразующего белка (порина), проницаема для определенных молекул.
Внутренняя митохондриальная мембрана имеет в несколько раз большую площадь поверхности, чем наружная мембрана. Ее поверхность значительно увеличивают кристы, ориентированные к центру органеллы. Кристы могут различаться по количеству, размеру и форме, при этом они имеют частицы, прикрепленные к ним с помощью коротких ручек. Эти частицы содержат АТФ-синтазы, ферментный комплекс, участвующий в синтезе АТФ.
Матрикс заполняет внутреннюю часть митохондрий и представляет собой смесь нескольких сотен ферментов, которые преобразуют продукты метаболизма углеводов, липидов и белков через цикл Кребса в углекислый газ и воду с выделением энергии в виде молекул АТФ. В этом процессе электроны переносятся по дыхательной электронной цепи, и происходит синтез высокоэнергетического фосфатного соединения, АТФ (окислительное фосфорилирование).
Читайте также:
